49 Words Between Brains
Language, Lateralization, and the Speaking Brain
49.1 Action aimed at another brain
Imagine a forager returning to camp without food. No one followed her across the ridge. No one else saw the tracks, the storm-damaged trees, or the herd moving through the valley beyond them. Yet she can describe where she went, explain why the hunt failed, warn that a river has risen, propose another route for tomorrow, and tell someone which tools to bring. She can correct a misunderstanding before anyone leaves. She can teach a child who has never seen the place. She can refer to an animal that is absent, dead, hypothetical, or invented.
Nothing in the pressure wave leaving her mouth resembles a river, a valley, an animal, or tomorrow.
Communication did not begin with humans. A honey bee can return to the hive and signal the direction and distance of food. Vervet calls alter the behavior of listeners in predator-specific ways. Songbirds learn elaborate vocal sequences from other birds. Primates coordinate voice, face, gaze, posture, and gesture. These systems should cure us of the idea that language appeared in an otherwise silent animal world.
Human language is nevertheless unusual. Learned symbols can refer beyond the present place and moment. A finite repertoire can be combined into messages that neither speaker nor listener has encountered before. Utterances are tailored to what a particular listener is expected to know. Failures are repaired while the interaction unfolds. Children reconstruct the local system without being explicitly taught most of its regularities, and the system they reconstruct has already been altered by generations of earlier users.
Language is therefore not merely a channel through which one brain reports an already completed thought. It is a way for one organism to alter the options, expectations, and actions of another.
A warning redirects attention and prepares the body. A request recruits another person’s movements. An explanation changes what the listener can predict. A promise reorganizes expectations about the future. A rule coordinates people who may never meet. A story allows an event experienced or imagined by one person to influence people separated from that person by distance, years, or death.
Language is action aimed at another agent.
The word control will recur in this chapter, and it needs the same caution used throughout the book. Control does not mean domination or perfect command. In control theory, an action is selected because it is expected to move a variable toward a desired range, and feedback reveals whether it succeeded. A speaker tries to make something happen in a listener—attention, understanding, agreement, amusement, fear, movement—and the listener remains an autonomous control system with goals of their own. Communication is therefore coupled control, often cooperative and sometimes adversarial.
This is the architecture-first starting point. Language did not descend on a completed brain as a disembodied faculty. It uses evolved systems for hearing and vision, breathing and movement, sequencing and prediction, memory and valuation, joint attention and social learning. Those systems are reorganized during development inside a community already speaking or signing. The biological capacity and the cultural inheritance cannot be separated cleanly, because neither produces a language user alone.
This is also why language belongs in the final unit. Chapter 48 argued that human adaptation increasingly resides in networks: calories, care, risk, and knowledge are distributed among many people and across generations. Language is one of the principal channels that makes such distribution possible. It turns private observation into public guidance and lets knowledge persist beyond the brain that acquired it.
The familiar neuroscience story begins with Broca’s area, Wernicke’s area, the left hemisphere, and split-brain patients. All matter, and we will discuss them. But they are evidence about a system, not the problem the system evolved and develops to solve. Before asking where language is located, we should ask what language adds to older communication, which ancestral systems could be recruited, why speech became prominent without exhausting language, and how a culturally inherited practice becomes organized in a developing brain.
49.2 What language adds
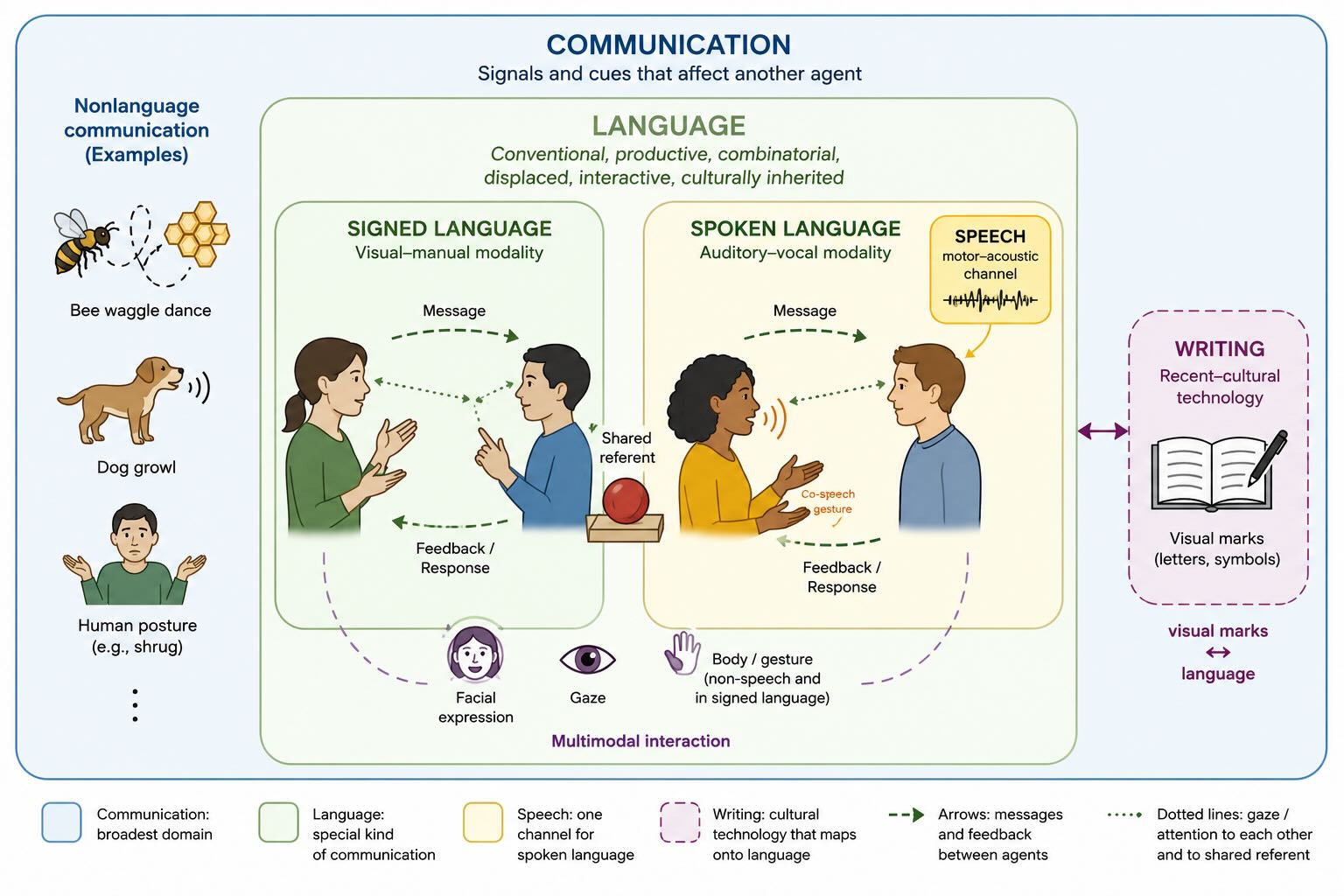
The word language is often used carelessly. It may refer to speech, communication, writing, vocabulary, grammar, or any organized signaling system. Those things overlap, but they are not identical.
Communication is the broadest category. A peacock’s tail, a bee’s dance, a dog’s growl, a cuttlefish’s changing skin, and the odor deposited by an ant can all alter the behavior of another animal. Some signals are deliberate and flexible; others are comparatively fixed. Some carry information because the signaler benefits from being understood; others reveal information despite the signaler’s interests. Communication is ancient and widespread.
Speech is one physical way of carrying language. It is an exquisitely controlled motor act in which breathing, phonation, tongue, lips, jaw, and auditory feedback are organized into rapidly changing acoustic patterns. Speech is biologically important because the vocal channel allows communication while the hands are carrying, making, grooming, or gesturing. It works in darkness and around partial visual obstruction. It can address several listeners at once. But speech is not language itself.
Signed languages make that distinction decisive. They possess conventional vocabularies, grammatical structure, productive combination, regional variation, historical change, poetry, ambiguity, humor, and all the other properties expected of language. Damage to the left hemisphere can produce aphasia in a signer even when the hands remain capable of movement. The linguistic system is therefore not tied to sound, and its characteristic left-hemisphere bias cannot be explained simply by the acoustics of speech [@emmorey2021]. Humans communicate multimodally: voice, hands, face, gaze, posture, and shared objects normally work together rather than competing to be the one true channel [@levinsonholler2014].
Writing is newer still. No human population needed to read in order for Homo sapiens to evolve, and many spoken languages have never had a writing system. Reading must therefore recruit neural machinery that evolved for other purposes. It is a cultural technology installed into a plastic visual and language system. We will return to that installation when we consider alexia.
What, then, is distinctive about language? No single list settles the matter, but several capacities become especially important when they occur together.
Words and signs are largely conventional: nothing about the sound river looks or sounds like flowing water. Speakers inherit an agreement—usually without realizing it—that this pattern will be treated as referring to that kind of thing. Language is combinatorial: a limited stock of elements can be assembled into messages that have never been produced before. It is displaced: it can refer beyond the present place and moment. It is productive: speakers do not merely choose from a fixed catalogue of calls. It is interactive: listeners signal confusion, speakers revise, and both participants use context to infer what was intended rather than merely decode what was said. And it is culturally inherited: the particular words, sounds, constructions, and conversational expectations available to a child are supplied by other people.
No child inherits English, Turkish, American Sign Language, or Yoruba in the genome. The genes build a learner. The group supplies a language.
This distinction matters because language is simultaneously biological and cultural. A child is prepared to learn language with striking speed and reliability despite receiving finite, variable, and often incomplete examples. Yet the child must encounter speakers or signers. The system that emerges is neither written in the genes nor invented by the child. It is reconstructed during development from the behavior of other people, and the language being reconstructed has itself been modified by generations of earlier learners.
The infant’s auditory system makes the loop visible. Early in life, infants discriminate speech contrasts that their community may not use. During the first year, sensitivity becomes increasingly tuned to the distinctions that organize the local language [@werkertees1984]. This is often described as perceptual narrowing, but narrowing can sound like simple loss. It is better understood as specialization. The nervous system becomes a more efficient detector of the boundaries that matter in the surrounding linguistic niche. Other brains train the infant’s auditory cortex to hear the categories those other brains use.
Language thus becomes part of development in the same sense that a nest, hive, school, or road becomes part of development. It is an inherited feature of the environment that organizes the nervous system growing within it.
49.3 A mosaic, not a miracle
The fossil record contains no preserved sentences. The soft tissues of the larynx and tongue rarely fossilize, and the anatomy of an endocast cannot tell us whether its owner could discuss a hunt, tell a joke, or embed one clause inside another. Language also leaves no single behavioral signature in stone. A carefully made tool may imply teaching, imitation, planning, or shared conventions, but it does not prove that the maker possessed modern syntax. Symbolic objects and long-distance trade are informative, but by the time they become abundant, much of the relevant biological evolution may already have occurred.
This uncertainty invites dramatic origin stories. A mutation produces grammar. A descended larynx produces speech. Mirror neurons produce meaning. A larger Broca’s area produces syntax. A social brain produces gossip. Each story captures something interesting. None is sufficient.
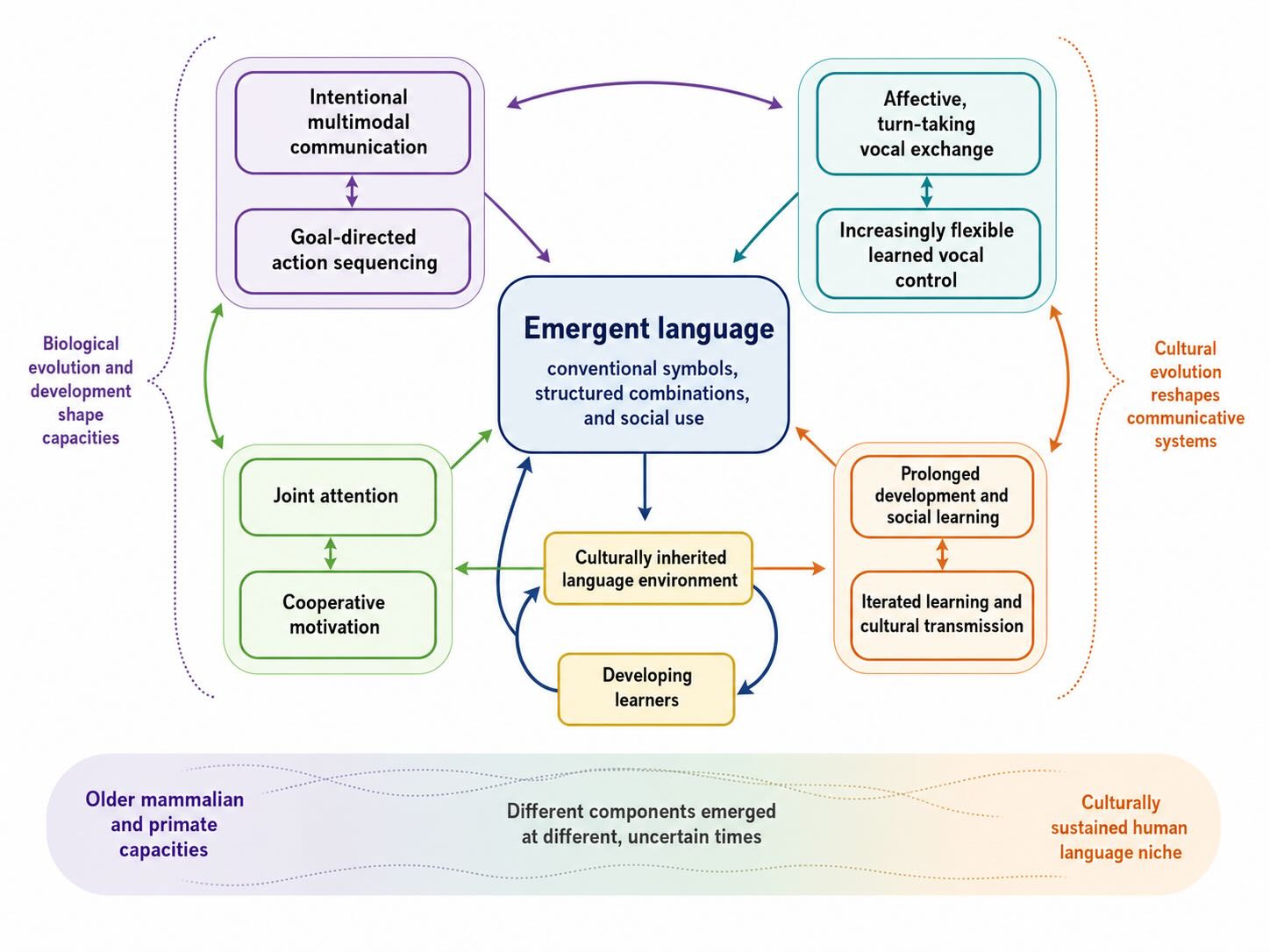
The reason is that language is not one adaptation. It is a coalition of capacities that have different evolutionary histories. Voluntary control of breathing and vocalization, rapid auditory analysis, learned mapping between signals and meanings, sequencing of actions, memory for words, sensitivity to other agents, shared attention, imitation, turn-taking, motivation to inform, and years of social learning did not need to appear at once. Many have recognizable precursors in other animals. What is unusual is their integration, scale, developmental reliability, and cultural amplification.
This changes the evolutionary question. Instead of asking, “When did language appear?” as though a switch were thrown, we can ask which components were already present in ancestral primates, which were modified in the hominin lineage, which emerged through interactions among components, and which properties belong partly to languages themselves as culturally evolving systems.
A mosaic account also prevents a common mistake. The fact that a component is found in another species does not make human language ordinary. Birds have vocal learning, apes use flexible gestures, monkeys sequence calls, and many social animals track what others are attending to. Conversely, the fact that no nonhuman animal combines these capacities as humans do does not require a miraculous new organ. Evolution often produces a qualitatively new system by changing the coordination among old parts.
The discontinuity may be real even when the components are continuous.
49.4 Several origin stories
There is no shortage of hypotheses about the evolution of language. They are often presented as rivals, but many address different parts of the problem. One asks where intentional symbols came from. Another asks how vocal control improved. Another asks why individuals were motivated to share information. Another asks how grammatical structure became learnable and stable. A sensible first draft of the history may therefore require several stories at once.
49.4.1 From hand to mouth
Michael Corballis made the gesture-first argument especially vivid. Nonhuman primates have relatively flexible control over their hands and arms, while many of their vocal calls are closely tied to emotional states. An ape can intentionally extend a hand, point the body, reach, beg, threaten, or elaborate a gesture when a recipient fails to respond. Manual action is also visible to the actor. A movement made by someone else can be related to movements in one’s own repertoire. This makes gesture a plausible early medium for intentional communication [@corballis2002].
The idea gains force from three observations. First, human gesture remains deeply integrated with speech. We move our hands even when gesture is not required to identify the referent, and listeners use those movements. Second, when people who share no language are asked to create a communication system from scratch, gesture often proves more intelligible than nonlinguistic vocalization, and repeated interaction rapidly conventionalizes the gestures [@fay2014; @fay2022]. Third, signed languages demonstrate that the visual-manual channel can carry a complete linguistic system rather than a pantomimed supplement to speech.
The gesture-first proposal also fits an appealing neural story. Monkey premotor area F5 contains neurons active during some goal-directed actions and while observing related actions performed by another individual. A roughly corresponding region in the human inferior frontal cortex includes what is traditionally called Broca’s area. Michael Arbib and others have proposed that a system originally involved in recognizing and organizing actions could have been elaborated into a system for pantomime, conventional gesture, and eventually language [@arbib2005]. On this view, the route to language passes through a circuit for doing and seeing actions rather than through a wholly new symbolic organ.
There are good reasons not to stop there. Human communication is normally multimodal, and vocalization did not wait outside history until gesture had finished building language. Primates use voices, faces, gaze, posture, and hands together. The mouth and hand are tightly coupled in feeding, tool use, infant interaction, and communication. A better version of the hypothesis is therefore not that a silent gestural language was completed and later transferred to the mouth, but that increasingly intentional multimodal communication gave gesture an early advantage while vocal control gradually expanded [@kendon2017; @levinsonholler2014].
The hand may have helped language begin without ever surrendering the conversation.
49.4.2 The voice before words
A different family of hypotheses begins with the ancient vocal system. Mammalian calls already regulate distance, affiliation, alarm, aggression, mating, and care. Primate vocal exchanges also have timing. Individuals alternate, respond, and sometimes adjust calls to the social context. Human conversation retains an astonishingly fast turn-taking rhythm: listeners begin preparing responses before a speaker has finished, using prediction and shared conventions to keep gaps brief [@levinson2016]. That conversational machinery may be older than words.
Darwin suspected that a music-like protolanguage preceded fully articulated speech. More recent musical protolanguage proposals emphasize pitch, rhythm, emotional prosody, synchrony, and socially learned vocal display [@fitch2006]. A melodic vocal system could coordinate groups, strengthen bonds, regulate infants, and communicate affect before it carried a large stock of arbitrary words. Anyone who has soothed an infant knows that voice can act on another nervous system before either participant exchanges propositions.
This hypothesis explains why language still rides on a richly expressive acoustic carrier. The same sentence can be reassurance, disbelief, warning, sarcasm, or threat depending on timing and prosody. Right-hemisphere and subcortical systems contribute importantly to these dimensions even when the left hemisphere is carrying most of the words. Voice is not a neutral pipe through which language passes. It is an older social control system that language colonized.
The difficulty is explaining the transition from graded, affective vocal patterns to stable symbols and compositional structure. A melody can coordinate and move a group without specifying who did what to whom. Musicality may have prepared timing, vocal learning, and social motivation, but it does not by itself produce a lexicon or grammar.
49.4.3 A tool for cooperative lives
A third set of hypotheses asks not which channel came first but why selection would favor such an expensive communication system. Human beings depend on unusually dense networks of cooperation. Food is shared, infants are provisioned by more than a parental pair, skills are divided among specialists, and reputations matter. In that setting, information about food, danger, competence, obligation, kinship, and alliance has direct biological value.
Robin Dunbar proposed that language functioned partly as vocal grooming. Physical grooming stabilizes relationships in many primates, but it is time-consuming and usually limited to one partner at a time. Vocal interaction can reach several partners while leaving the hands available, allowing larger groups to maintain affiliation and monitor relationships [@dunbar1993]. The proposal is sometimes caricatured as “language evolved for gossip,” but its serious point is that talk performs social work. Much everyday conversation is not a technical transfer of facts. It establishes attention, trust, status, inclusion, and common ground.
Michael Tomasello and colleagues place greater emphasis on shared intentionality: the capacity and motivation to pursue joint goals, share attention, and understand communication as an attempt to alter what another person knows or does [@tomasello2005]. A point is informative only when a recipient understands not merely the direction of the finger but why the other person is directing attention. Human infants enter routines of joint attention long before they command grammar. They follow gaze, offer objects, seek confirmation, and use adults as partners in a shared problem.
This social-pragmatic account fits the argument of the previous chapter. An expensive, slow-developing offspring must learn from a network of caregivers and experts. A group whose members can teach routes, foods, tools, norms, and dangers can support individuals who are individually less self-sufficient but collectively more capable. Language does not merely make a large brain useful. It helps make a large brain affordable by allowing knowledge acquired by one person to guide many others.
The limitation is that social need alone cannot explain the form of language. Baboons, elephants, dolphins, and wolves also live consequential social lives. Cooperative breeding and joint action create a selection pressure and a developmental niche; they do not specify how symbols become organized into sentences.
49.4.4 The language that learned the learner
The fourth story shifts attention from the evolution of brains to the evolution of languages. A language must repeatedly pass through a severe bottleneck: it has to be learned by new individuals from finite and imperfect samples. Forms that are impossible to remember, difficult to distinguish, or opaque to infer are less likely to survive transmission. Languages therefore adapt culturally to the learning and use biases of human brains.
Experiments using iterated learning make the principle visible. One participant learns an artificial signaling system and produces it for the next participant, whose output becomes the input to another. As the system moves down the chain, idiosyncratic details often disappear and more regular, compressible, or compositional patterns emerge. No participant designs the final structure. It is an unintended product of repeated learning and use [@kirby2008].
This does not mean biology is irrelevant. Cultural systems can only explore forms that human bodies and brains can perceive, learn, and produce. But it does mean that some apparent design in language may belong to the long interaction between learners rather than to a genetically specified grammar. Biology shapes the channel; cultural transmission shapes what reliably travels through it; the resulting languages alter the developmental environment of the next generation [@kirby2017].
The loop is precisely the one introduced in the overview. Brains make languages. Languages reorganize developing brains. Languages that are easier for those brains to learn and use are more likely to persist. The culturally inherited system becomes a selective and developmental niche.
The language learned the learner.
49.4.5 No single winner
Gesture-first, musical protolanguage, vocal grooming, shared intentionality, and cultural evolution are not interchangeable, but neither are they mutually exclusive. Gesture may have supported deliberate iconic reference. Vocal exchange may have supplied timing, affect, and a channel that freed the hands. Cooperative social life may have made information sharing worth its costs. Iterated transmission may have transformed unstable inventions into conventional and increasingly structured systems. Biological evolution could then favor individuals better able to acquire and use the cultural system already surrounding them.
The honest conclusion is not that we know which hypothesis won. It is that language probably has no single point of origin to award.
Mirror neurons became one of neuroscience’s most rapidly expanding metaphors. A neuron fires when a monkey grasps an object and when the monkey sees another animal perform a related action; perhaps, the argument went, this is the neural basis of understanding action, imitation, empathy, culture, and language.
The observation is important. Perception and action are not sealed modules, and circuits used to organize one’s own behavior can contribute to predicting or interpreting the behavior of others. The anatomical relationship between monkey area F5 and human inferior frontal cortex makes an action-to-communication pathway evolutionarily plausible [@arbib2005].
But plausibility is not an explanation. Mirror responses do not by themselves provide conventional symbols, shared intentions, a lexicon, syntax, displacement, or the motivation to inform. An animal can map a seen grasp onto its own motor system without understanding a sentence, and humans can understand actions they cannot perform. The mirror system hypothesis is best treated as a proposal about one useful piece of ancestral machinery—not as the moment language was discovered hiding inside premotor cortex.
49.5 Why speech became dominant in many hearing communities
Even if intentional communication began partly in the hands, speech became the principal linguistic channel in many hearing communities. That outcome is not mysterious. The voice can operate while the hands are occupied with tools, infants, food, or locomotion. It reaches listeners who are not looking. It works at night, through vegetation, and across modest distances. A speaker can address several people at once, and the acoustic stream can be produced rapidly enough to carry a large number of distinctions in a short time.
Speech also allows two channels to cooperate. A person can name an object while pointing to it, describe an action while demonstrating it, or use the face and voice to mark whether an utterance is serious, playful, doubtful, or urgent. Gesture did not lose when speech became dominant. Speech made gesture more powerful by giving it a synchronized symbolic partner.
The transition nevertheless required unusual motor control. Most primates vocalize, but their calls are more tightly coupled to motivational and emotional states than human speech. A frightened monkey does not calmly decide which alarm call would best support a hypothetical example. Humans retained ancient pathways for crying, laughing, screaming, and affective vocalization while developing far greater cortical control over respiration, laryngeal activity, articulation, and learned vocal sequences [@nieder2020].
This is one reason speech production is a more compelling focus for hemispheric specialization than many of the broad claims made about “left-brained” and “right-brained” people. Speaking requires an extraordinary sequence of precisely timed actions. Sounds overlap. The tongue may already be moving toward the next consonant while the current vowel is still being voiced. The respiratory system must provide a controlled stream of air; the larynx converts it into a source signal; the vocal tract reshapes that signal; auditory and somatosensory feedback update the plan while it is unfolding. The output is not a chain of separate muscular commands but a continuously controlled trajectory.
The left hemisphere, in most people, is especially important for organizing that trajectory. But it does not do so alone, and it did not evolve a new set of muscles for language. It recruited a hierarchy of older motor systems.
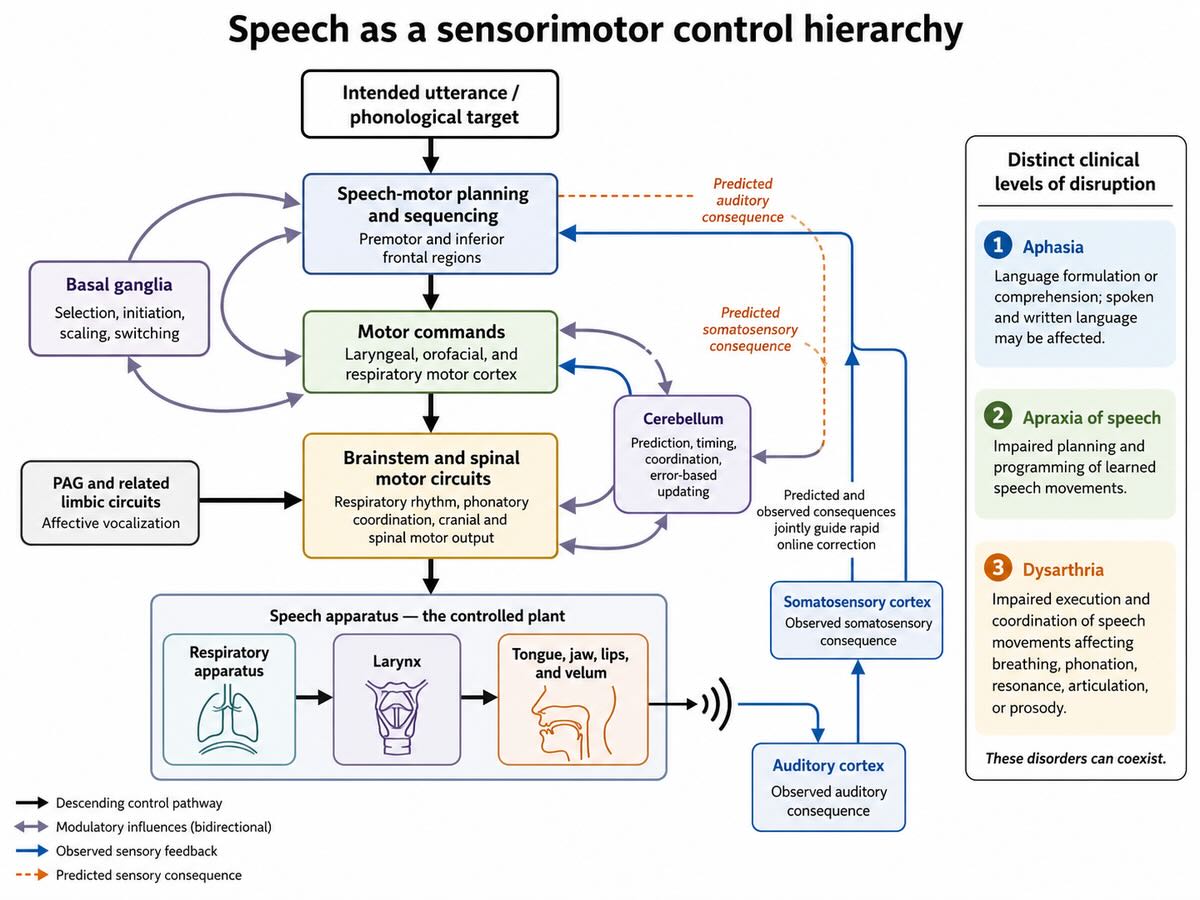
49.6 The machinery of speech
Speech begins below cortex. Brainstem circuits regulate breathing and coordinate vocal and orofacial musculature. The periaqueductal gray and related subcortical systems contribute to affective vocalization. Motor cortex supplies descending control over the face, tongue, larynx, and respiratory effectors. Premotor and inferior frontal regions organize sequences and transform intended sound patterns into articulatory plans. Auditory cortex analyzes the resulting sound, while somatosensory cortex reports the configuration and contact of the articulators. The basal ganglia contribute to selection, initiation, scaling, and switching; the cerebellum contributes prediction, timing, and error correction.
Calling all of this a “language module” would hide the important biology. Speech is a special use of systems for movement, sensation, prediction, and control.
The laryngeal motor cortex is particularly revealing. Human speech depends on flexible cortical control of respiration, phonation, and articulation, including descending access to brainstem motor systems. The exact homologies of laryngeal representations and the degree to which human corticobulbar pathways are categorically different from those of other primates remain debated. Nonhuman primate calls are not wholly inflexible, and human affective vocalizations still depend strongly on older subcortical routes. The defensible claim is that the human lineage greatly expanded the voluntary, learned, and rapidly sequenced cortical control required for speech [@simonyan2011; @hickok2017]. A lowered adult larynx contributes to vocal-tract acoustics, but it is neither uniquely human nor sufficient for language. The crucial change was an integrated control system, not one piece of plumbing.
Auditory feedback closes the loop. People alter articulation when the sound of their own voice is experimentally shifted. Hearing loss early in life affects the development of spoken articulation unless another linguistic channel is available. Yet the feedback is not slow correction after each sound. Speakers predict the sensory consequences of a motor command and compare incoming sound with the predicted result. The same control logic encountered throughout this book—prediction, action, feedback, correction—operates at the scale of tens of milliseconds while a sentence is produced.
The listener faces a complementary prediction problem. Ordinary speech does not insert a clean silence at every word boundary. Sounds are coarticulated, syllables vary with speaker and context, and a phrase such as what do you mean may form one continuous acoustic envelope. Auditory cortex supplies features, not a transcript. The brain segments the stream by combining learned sound patterns, probabilities, lexical candidates, syntax, and context. We hear words partly because years inside a language have taught the system what sequences are likely to count as words.
The basal ganglia provide another link to earlier units. A bilingual speaker must select one language while suppressing another, maintain that selection, and switch when the context changes. Damage involving the left caudate can disturb this control, sometimes producing unwanted switching or unequal access to languages. Parkinson’s disease can reduce speech volume, initiation, and prosodic variation. These effects do not make the basal ganglia a language center. They show that selecting a word or language is still a form of action selection.
The cerebellum similarly contributes. Its predictive machinery helps coordinate the timing and sensory consequences of speech. Cerebellar damage can produce dysarthria—poorly timed, imprecise articulation—even when the person knows exactly what they want to say. A fluent sentence therefore depends on structures that traditional cortical maps leave uncolored.
This distinction becomes clinically important. Aphasia is an impairment of language: selecting words, constructing or understanding sentences, mapping signals to meanings, reading, or writing. Dysarthria is an impairment of the muscular execution of speech. Apraxia of speech is a disturbance of planning and programming the articulatory movements required for speech. They can coexist, especially after a large stroke, but they are not the same problem. A person can possess language but be unable to articulate it clearly, or articulate fluent sound while failing to stabilize shared meaning.
The most famous genetic story begins with the KE family, many of whose affected members have severe difficulties with speech articulation, orofacial praxis, grammatical morphology, and aspects of language comprehension. The pattern followed an autosomal dominant inheritance, and investigators identified a mutation in FOXP2, a gene encoding a transcription factor that regulates many other genes [@lai2001].
The finding was important because it connected a molecular change to the development of neural systems required for learned vocal behavior. It did not identify a gene containing grammar.
FOXP2 is evolutionarily ancient and expressed in circuits involved in motor learning, including cortex, basal ganglia, and cerebellum. Related genes occur across vertebrates. In zebra finches, reducing FoxP2 expression in Area X, a basal-ganglia-like nucleus important for song learning, leads to incomplete and inaccurate imitation rather than simply abolishing song [@haesler2007]. The result points to a conserved role in learning and sequencing vocal motor behavior.
The human protein differs at a small number of sites from that of chimpanzees, but the derived coding variant is also found in Neanderthals [@krause2007]. Modern human speech therefore did not appear when a uniquely human “language gene” switched on. FOXP2 is one component in a large developmental regulatory network, and the KE-family phenotype includes motor as well as linguistic difficulties [@fisher2009].
The useful lesson is broader. Genes do not encode words or sentences. They influence the development and plasticity of circuits that make certain kinds of learning and control possible. Language emerges when those circuits develop inside a community already using language.
49.7 One language system, unequal hemispheric roles
The cerebral hemispheres differ systematically in anatomy and function. The planum temporale is often larger on the left, and population-level torque and other anatomical asymmetries are well established. The clearest functional asymmetry is language. In most people, language is strongly left-hemisphere dominant. Words flashed briefly in the right visual field are often reported more efficiently than words flashed in the left, and spoken verbal material can produce a modest right-ear advantage. The strongest evidence comes from lesions, electrical cortical stimulation, and temporary hemispheric inactivation, with functional imaging providing convergent evidence. Disrupting the left language network is much more likely to impair speech production, naming, phonological processing, and sentence construction than comparable disruption of the right hemisphere [@toga2003; @hamberger2011; @corballis2014].
The Wada test made this asymmetry clinically unmistakable before functional MRI became widely available. Amobarbital injected into one internal carotid artery temporarily suppresses function in much of that artery’s hemispheric territory. In a patient with typical left-hemisphere language dominance, left-sided injection can produce speech arrest and marked failures of naming or other language functions; right-sided injection usually does not. The procedure is invasive, its vascular distribution is imperfect, and functional MRI has replaced it in many presurgical evaluations. Nevertheless, its central lesson remains: for most people, the hemispheres are not interchangeable with respect to language [@baxendale2009; @hamberger2011].
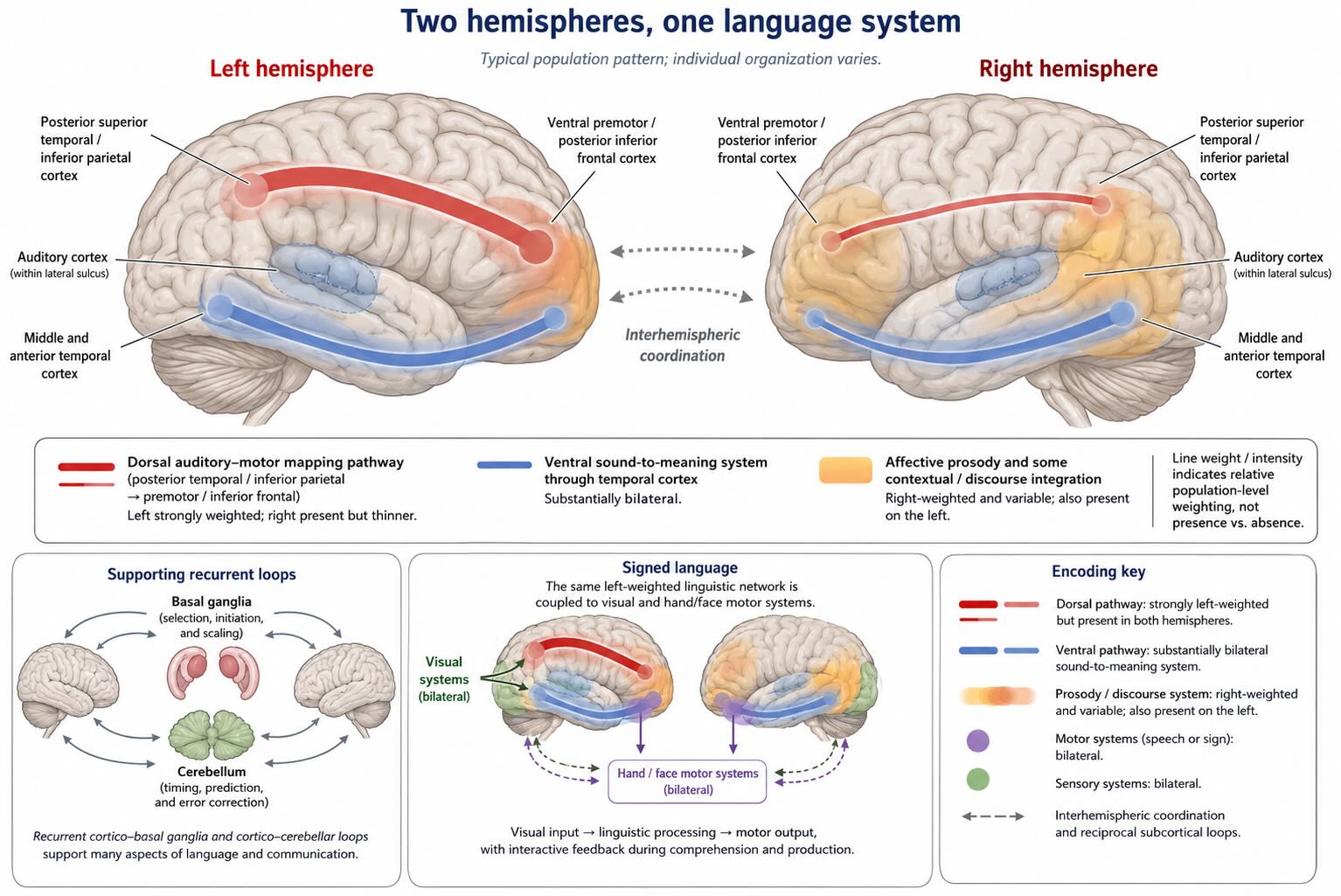
Left dominance does not mean left exclusivity. The left hemisphere is dominant for speech production, phonological sequencing, rapid auditory–motor mapping, lexical retrieval, and many aspects of grammatical processing. The ventral pathways that connect speech sounds with lexical and conceptual meaning are more bilaterally organized, although bilateral representation does not imply equal contribution. The right hemisphere makes complementary contributions to emotional prosody and to aspects of discourse and pragmatic interpretation. Right-hemisphere damage can therefore leave words and sentence structure relatively intact while impairing the production or interpretation of emotional tone. These functions are less uniformly lateralized than speech production, but their variability does not erase the much stronger leftward asymmetry of the core language network [@hickok2007; @ferre2011].
Signed languages show that this organization is not an artifact of hearing speech or controlling the vocal tract. Signers recruit a predominantly left-lateralized language network, and left-hemisphere damage can produce aphasia in sign just as it can in speech. Signing also engages modality-specific visual and motor systems, but the dominance of the core linguistic network remains leftward. Language lateralization is therefore a property of language organization across modalities, not merely a consequence of auditory input, vocal production, or superior control of the right hand [@emmorey2021].
Handedness shifts the probability of language dominance; it does not determine it. Most right-handed people are left-dominant for language. Most left-handed people are also left-dominant, although bilateral or right-hemisphere organization is more common among left-handers. The hand that holds the pencil is therefore a clue about language organization, not a map of it [@knecht2000].
None of this supports the popular division of people into globally “left-brained” analytical types and “right-brained” creative types. Brain functions can be lateralized without whole persons being lateralized in one direction across all functional networks. The myth is not that hemispheric specialization exists. The myth is that it defines personality [@nielsen2013; @corballis2014].
Why language became lateralized is less certain than the fact that it did. Proposed advantages include keeping tightly interacting operations close together, reducing conduction delays, and limiting interference between partially competing computations. Development may then amplify an initial asymmetry: a hemisphere that gains an early advantage receives more practice and develops stronger within-hemisphere connections. These are hypotheses about how specialization arises, not reasons to doubt that specialization exists [@corballis2014].
One plausible evolutionary interpretation is that language recruited older systems with different prior histories. Affective vocalization, social perception, and auditory scene analysis long predate human language and were already distributed across the hemispheres, often with their own asymmetries. In this view, human language developed much stronger leftward specialization for many rapidly ordered sensorimotor and combinatorial operations. Signed language shows, however, that this specialization cannot be explained by vocal control alone. Language did not move into an empty left hemisphere; it assembled a system across both hemispheres while assigning them markedly unequal roles [@corballis2014; @emmorey2021].
The accurate picture is therefore neither two separate minds nor two equivalent halves. It is one integrated language system with marked left-hemisphere speech dominance and complementary right-hemisphere contributions.
Lateralization is division of labor, not division into two people.
The classic split-brain experiments are among the clearest causal demonstrations in cognitive neuroscience. Patients with otherwise intractable epilepsy underwent commissurotomy or callosotomy to prevent seizure activity from spreading between the hemispheres. In the most completely disconnected cases, the corpus callosum was sectioned together with some or all of the smaller cerebral commissures. The patients were few and neurologically heterogeneous, and the exact surgical anatomy differed among them. Those qualifications limit generalization, but they do not diminish the principal result: after sufficiently complete disconnection, complex perception, memory, choice, and action could proceed in one hemisphere without the relevant information becoming available to the other [@gazzaniga1967; @volz2017; @dehaan2020].
When a word was flashed briefly to the right visual field, it reached the usually language-dominant left hemisphere and could be named. A word flashed to the left visual field reached the right hemisphere and often could not be spoken. Yet the left hand, controlled primarily by the right hemisphere, could select or draw the corresponding object. The right hemisphere had processed the stimulus well enough to guide an appropriate response; the speaking left hemisphere lacked access to that information. Split-brain research therefore separated three things that ordinarily seem inseparable: processing a stimulus, using it to guide behavior, and being able to explain verbally what happened.
Information could sometimes pass between the hemispheres by an external route even when it could no longer cross through the cerebral commissures. The right hemisphere might guide the left hand in drawing an object; once the drawing was visible in both visual fields, the speaking left hemisphere could identify it. The deficit was therefore not simply an absence of knowledge. It was a failure of one hemisphere to gain access to knowledge represented in the other.
The experiments also exposed a striking feature of causal explanation. In the classic chicken-claw and snow-scene task, the left hemisphere saw a chicken claw while the right hemisphere saw a snow-covered scene. The right hand selected a picture of a chicken, while the left hand selected a shovel. Asked to explain both choices, the speaking left hemisphere correctly related the chicken to the claw. Because it had not seen the snow scene, however, it explained the shovel as something used to clean the chicken shed. Rather than report that it did not know why the left hand had chosen the shovel, it constructed a coherent causal account from the information available to it [@gazzanigaLedoux1978; @gazzaniga2000; @volz2017]. Gazzaniga called this interpretive capacity The Interpreter. The name does not require a little storyteller located at a single point in the left hemisphere. It is better understood as a functional hypothesis: left-lateralized systems involved in language and inference organize accessible perceptions, actions, memories, and feelings into a causal narrative. When the relevant evidence is available, this capacity supports explanation, prediction, and understanding. When critical evidence is missing, the same drive for coherence can produce a plausible but incorrect account.
The chicken-and-shovel result was not the only evidence for this proposal. In other experiments, the left hemisphere showed a greater tendency to impose patterns on probabilistic events, infer causal relations, and incorporate the general meaning of an event into memory even at the cost of falsely recognizing something that had not occurred. The right hemisphere sometimes remained more closely tied to the information that had actually been presented. These findings do not establish a single, sharply bounded Interpreter module, but they support the broader idea that the language-dominant hemisphere is especially disposed to seek explanations and impose coherence [@phelps1992; @wolford2000; @roser2005].
The Interpreter also offers a useful, though incomplete, way to think about anosognosia. Anosognosia for hemiplegia can arise from failures of motor prediction, error monitoring, attention, representation of bodily state, and the updating of beliefs about one’s capacities. The Interpreter does not explain those initial failures. It may help explain what follows from them. When evidence of paralysis is unavailable or given too little weight, the patient’s established model—“my arm moves normally”—remains in place. The verbal system can then construct explanations that preserve that model despite contradictory events. On this account, confabulation is neither simple dishonesty nor an arbitrary response to questioning. It is causal coherence constructed from an impoverished evidence base [@orfei2007; @kirsch2021].
The relation between split-brain research and consciousness is deeper but less settled. Callosal disconnection can divide perceptual access and allow some decisions and actions to proceed independently, yet split-brain patients generally behave as single persons in everyday life and do not ordinarily report that their experience has divided in two. Available evidence does not determine whether complete disconnection produces two conscious subjects, one subject with divided access, or a partially unified organization that fits neither description [@dehaan2020]. The Interpreter is therefore not a complete explanation of consciousness. It is a serious proposal about one important feature of conscious human life: how a distributed brain produces the experienced coherence of a self with reasons, intentions, and a continuous history. Most of the systems that determine our behavior do not supply verbal reports of their computations. The speaking hemisphere receives their conclusions—the choice, the feeling, the movement, the preference—and constructs an account of why they occurred. Usually that account incorporates enough valid evidence to be useful. Split-brain experiments reveal what happens when the true cause of an action has been deliberately withheld from the system responsible for explaining it.
Methodological qualifications remain important. Patients differed in epilepsy history, surgical procedure, residual commissural connections, and opportunities for information to cross by indirect routes. Partial callosotomy should not be treated as equivalent to complete disconnection. These limitations define the boundaries of the evidence; they do not reduce the work to a neurological curiosity. Split-brain research showed that perception, knowledge, action, verbal report, and causal self-understanding can come apart—and that the apparent unity of the person may depend partly on the brain’s continuing interpretation of its own activity.
49.8 Reading a brain from its failures
Long before brain imaging, language entered neurology through catastrophe. The Edwin Smith papyrus describes head injuries and notes of one patient, with striking economy, that “he speaketh not to you.” In the nineteenth century, Marc Dax collected cases linking loss of speech with left-hemisphere injury. Paul Broca’s report of Leborgne—known as Tan because that was the syllable he most reliably produced—made the relationship famous. Carl Wernicke soon described a contrasting syndrome in which speech remained fluent while comprehension and meaningful word selection deteriorated.
These cases were powerful because language is otherwise invisible. A healthy speaker produces a sentence, and the cooperating machinery disappears into success. A lesion interrupts part of the system and exposes dependencies that ordinary behavior conceals.
Stroke supplied many of the crucial lesions. The middle cerebral artery carries blood to much of the lateral frontal, parietal, and temporal cortex. An embolus or local clot that blocks a left middle cerebral branch can deprive language-related cortex and white matter of oxygen and glucose. The resulting infarct may damage inferior frontal cortex, auditory association cortex, parietal regions, insula, basal ganglia, and the pathways among them. Because vascular territories do not respect textbook boundaries, real aphasias are often mixtures rather than clean demonstrations of one damaged region.
This is why the classical syndromes should be treated as recognizable patterns, not as rigid species of disorder. “Broca’s aphasia” and “Wernicke’s aphasia” remain useful clinical shorthand. They tell us something about fluency, comprehension, repetition, and likely lesion distribution. They do not mean that every patient has the same deficit or that a single cortical patch contains the lost function.
Aphasia also does not imply loss of intelligence, memory, emotion, or personhood. A patient may understand a situation, recognize people, plan an action, feel embarrassment, and know exactly what they wish to communicate while being unable to retrieve or organize the words. Another may produce long, confident utterances while failing to notice that the words no longer establish shared meaning. The neurological deficit changes access to language; it does not convert the patient into a simpler human being.
The clinical examination therefore asks several questions rather than simply “Can this person talk?” Is spontaneous speech fluent? Are phrases grammatically organized? Can the patient understand words and sentences? Name an object? Repeat a phrase? Read? Write? Follow a command? Does performance change when syntax becomes complex, when a word is unusual, or when a sentence has to be held in working memory? The profile matters more than the label.
49.9 Broca: when a sentence will not come
Broca’s patient Leborgne could understand ordinary speech better than he could produce it. He could move his tongue and face, respond emotionally, and communicate through gesture, yet propositional speech was reduced largely to tan. After death, Broca found extensive damage in the left frontal lobe. The preserved brain has since been scanned, revealing that the lesion extended well beyond the small patch now colored as Broca’s area in introductory diagrams [@dronkers2007].
The syndrome called Broca’s aphasia is usually described as nonfluent or effortful. Speech emerges slowly, with shortened phrases and long pauses. Function words and grammatical endings may be omitted, producing the famous “telegraphic” style: son—college—smart—good. Naming is difficult. Repetition is impaired. Patients often know that their speech is failing and work visibly to overcome it.
Some highly practiced or emotionally charged utterances can be easier than novel propositional speech. A patient who struggles to construct a sentence may produce a familiar greeting, count, sing part of a song, or swear with surprising fluency. These preserved islands are variable and should not be romanticized into a simple right-hemisphere cure. They do show that formulaic language, melody, affect, and newly assembled sentences draw differently on cortical and subcortical systems.
It is tempting to hear only a motor problem, but language is affected more deeply. Many patients have difficulty interpreting grammatical relationships when word order and real-world plausibility pull in different directions. The boy hit the girl is easy because the first noun is the likely agent. The boy was hit by the girl requires the listener to use grammatical structure to reverse that expectation. A patient who performs well in casual conversation may fail on precisely such sentences. Broca’s aphasia is therefore not simply paralysis of speech.
At the same time, nonfluent output can include several overlapping disorders. A large left frontal stroke may cause right facial and limb weakness, dysarthria, apraxia of speech, impaired working memory, and aphasia together. The clinician must determine whether a sound is wrong because the muscles are weak, the articulatory sequence cannot be programmed, the word cannot be retrieved, or the sentence cannot be grammatically assembled. The patient’s struggle may arise at more than one level.
Direct cortical stimulation makes one part of the syndrome vividly concrete. During neurosurgery, a patient can be asked to count, name pictures, or read while brief electrical stimulation is delivered to the exposed cortex. Stimulation of a site in left inferior frontal or adjacent premotor cortex may produce speech arrest: the patient stops mid-utterance, yet can still protrude the tongue or move the lips on command. When stimulation ends, speech returns. The result does not prove that a sentence is stored at that point. It shows that the site is a necessary node, under those conditions, in the network converting a linguistic plan into spoken action.
I have shown such maps in class from patients studied before surgery. The anterior language sites could be surprisingly compact: stimulation interrupted counting or reading without producing ordinary face movement. A few centimeters posterior, stimulation moved the mouth or produced somatic sensation. Farther back, the effects became more varied—naming errors, paraphasias, failed sentence completion, or inability to follow a command. The map looked less like two circled organs than like a network whose nodes could be interrupted in different ways.
Modern lesion studies support that impression. Persistent speech production deficits after stroke are not explained by Broca’s area alone. Damage to surrounding frontal cortex, premotor regions, insula, underlying white matter, and posterior language regions contributes importantly. Some patients with injury confined largely to Broca’s area recover fluent speech; many with chronic Broca-like aphasia have much larger lesions [@gajardovidal2021; @fridriksson2018].
Broca was not wrong. Injury in the left inferior frontal region can profoundly disrupt language, and the region participates in articulation, sequencing, selection, and sentence-level processing. The error came later, when a lesion association was turned into a container: syntax or speech production was imagined to reside inside an anatomically neat box.
The box is useful on a first map. It is not where the explanation ends.
49.11 Other ways the network can fail
The classical contrast between Broca and Wernicke is memorable because it creates two poles: effortful output with comparatively better comprehension, and fluent output with impaired comprehension and unstable meaning. Many patients occupy neither pole.
49.11.1 Anomia: the missing name
Anomia is difficulty retrieving words, especially the names of people and objects. It appears in many aphasias and can dominate the residual deficit after other abilities recover. The person knows what the object is and may describe its use, shape, location, or associated actions, yet the name remains inaccessible. A cookie becomes “the thing that is not good for you but you love.” A saw becomes “the thing in the workshop that cuts the wood.” This circumlocution is not evidence that the concept has vanished. It is often an intelligent route around a blocked lexical path.
Word retrieval is not a single lookup. Seeing an object activates visual representations, conceptual knowledge, possible names, their sounds, and an articulatory plan. Competition must be resolved and the selected form maintained long enough to speak. Damage at different points produces different errors. A semantic paraphasia such as fork for spoon suggests access to the right conceptual neighborhood but selection of the wrong member. A phonemic paraphasia changes or rearranges sounds. A neologism produces a form not recognized as a word. The error tells us something about where the trajectory departed from its target.
All healthy speakers experience occasional tip-of-the-tongue states, especially for proper names. Aphasic anomia is not merely more of the same inconvenience. It can make every sentence a search task and every conversation a public test.
49.11.2 Conduction aphasia: hearing without easy repetition
In conduction aphasia, spontaneous speech and comprehension may be relatively good while repetition is disproportionately impaired. A patient understands a phrase, intends to reproduce it, and then generates phonemic errors while repeatedly trying to correct them. The preserved awareness of error can be as striking as the errors themselves.
Norman Geschwind interpreted the syndrome as a disconnection between a posterior comprehension center and an anterior production center, carried by the arcuate fasciculus. The model had enormous pedagogical power: Wernicke’s area understands, Broca’s area speaks, and a severed wire prevents one from handing the message to the other.
The modern account is both less simple and more interesting. Repetition depends on a dorsal auditory-motor pathway that maps heard sound onto articulatory representations and temporarily maintains phonological sequences. The arcuate fasciculus is part of that system, but conduction aphasia can follow cortical as well as white matter damage, and the relevant network includes posterior temporal and inferior parietal regions involved in phonological working memory [@buchsbaum2011]. The syndrome is therefore a failure of sensorimotor transformation and maintenance, not merely a cut telephone line between two independent word centers.
49.11.3 Global aphasia and the size of the lesion
A large left middle cerebral artery stroke can produce global aphasia, with severe impairment of fluent production, comprehension, naming, and repetition. Global aphasia is a reminder that vascular lesions often erase the boundaries that teaching diagrams draw. The patient may also have right hemiplegia, sensory loss, visual-field deficits, and apraxia. Over time, the profile may change as edema resolves, tissue recovers, and surviving networks reorganize.
The label describes severity across domains; it does not predict a fixed endpoint. Some patients recover useful comprehension or formulaic speech. Others remain profoundly impaired. Age, lesion location and volume, vascular health, therapy, social support, and the integrity of connected tissue all matter.
49.11.4 Pure word deafness: sound without speech
A rarer disorder, pure word deafness, further separates language from hearing. A patient may detect sounds, identify environmental noises, and sometimes appreciate music while spoken words arrive as unintelligible acoustic events. Reading and speaking can remain relatively preserved. The deficit can follow bilateral superior temporal damage or a strategically placed left temporal lesion that disrupts access from auditory analysis to phonological representations.
The name is imperfect—the disorder is rarely pure—but the dissociation is conceptually useful. The ear and primary auditory pathways can deliver sound while the learned categories that make the sound speech fail to stabilize. A spoken word is not present in the waveform waiting to be extracted. It is an interpretation constructed by a trained auditory-language system.

49.12 Reading is cultural recycling
Speech is ancient enough to have influenced human biological evolution. Reading is not. The oldest writing systems are only several thousand years old, far too recent for natural selection to have designed a cortical organ specifically for printed words. Yet literate adults often show a reproducible region in left ventral occipitotemporal cortex that responds strongly to familiar letter strings and written words—the region commonly called the visual word form area.
This is not a contradiction. A culturally invented skill can become neurally specialized during development by repeatedly recruiting a circuit whose prior organization makes it suitable. The ventral visual pathway already transforms lines, contours, and object parts into increasingly stable representations. Language networks already represent sounds, words, and meanings. Learning to read creates efficient mappings between them. Dehaene and colleagues call this neuronal recycling: culture occupies cortical territory constrained by older functions and reshapes it without beginning from blank tissue [@dehaene2010; @dehaenecohen2011].
Our early intracranial recordings made the scale of this specialization memorable. Electrodes only a centimeter or two apart in inferior temporal cortex could show very different responses. One site responded strongly to letter strings or names; a nearby site responded strongly to faces. Functional imaging localized corresponding patches in neurologically intact participants. The cortex had not evolved for the name Bill Clinton or for the printed alphabet. Experience had tuned neighboring parts of an older recognition system to culturally important categories [@nobre1994].
Damage reveals the pathway. In pure alexia, or alexia without agraphia, a patient may write a sentence and then be unable to read it. Reading becomes slow and letter-by-letter. Some patients trace letters with a finger or pronounce them individually, converting a difficult visual route into a partially preserved motor or auditory route. The paradox disappears once reading and writing are understood as different paths through a network. Writing can be driven from language and motor systems, while visual access from print back into the language system is disrupted.
Pure alexia often follows damage to left occipitotemporal cortex, sometimes combined with injury to the splenium of the corpus callosum. A left visual lesion can remove direct visual input to the left reading network; splenial damage can prevent intact right occipital cortex from transferring visual information across the midline. The syndrome can therefore reflect damage to a specialized visual region, disconnection of that region from visual input, or both.
Alexia with agraphia usually involves a broader left temporoparietal lesion and impairs both reading and writing. Again, the syndrome should not be assigned to a single “reading center.” Written language recruits visual recognition, phonology, semantics, orthographic memory, attention, eye movements, and hand control. Different lesions remove different combinations.
Reading provides one of the clearest examples in the book of culture building a brain. A child enters a world containing marks that earlier humans invented. Years of instruction and practice alter visual cortex, strengthen pathways to spoken language, change phonological awareness, and create an apparently specialized cortical response. The cultural niche does not merely provide content to an unchanged brain. It reorganizes the machinery that encounters the content.
49.13 From centers to streams
Broca and Wernicke remain relevant because the syndromes they described are real and because the anterior-posterior contrast captures something important. Frontal systems contribute strongly to speech planning, sequencing, selection, and controlled combination. Posterior temporal systems contribute strongly to speech analysis, lexical access, and the representation of sound patterns. White matter connections allow these systems to cooperate.
The problem is the line usually drawn between two circles.
Contemporary models describe multiple interacting pathways. A ventral stream maps speech sounds onto lexical and conceptual representations—roughly, sound to meaning. It runs through superior and middle temporal regions and connects forward into temporal and frontal cortex. It is substantially bilateral, although the hemispheres need not contribute identically. A dorsal stream maps speech sounds onto articulatory and motor representations—roughly, sound to action. It links posterior temporal and inferior parietal regions with premotor and inferior frontal cortex and is more strongly left-lateralized [@hickokpoeppel2007; @saur2008].
The labels are useful if we remember that each stream contains several routes and that meaning is not a parcel delivered to a destination. The ventral stream supports increasingly abstract transformations between acoustic patterns, words, and concepts. The dorsal stream supports auditory-motor integration, phonological working memory, repetition, and learning to produce unfamiliar sound sequences. Frontal control systems select among competing words and constructions. Inferior parietal regions help maintain and transform phonological information. Anterior temporal cortex contributes to conceptual combination. The basal ganglia and cerebellum regulate selection and timing. The network changes with the task. Recent whole-brain intracranial recordings likewise find articulatory, acoustic, and semantic representations in both hemispheres, with more extensive and dynamic organization on the left [@verwoert2025].
Functional MRI visualizes this dependence on task with seductive clarity. In an early study from our laboratory, a bilingual speaker recorded matched sentences in English and Turkish, allowing listeners who understood English but not Turkish to hear closely matched acoustic streams. The contrast highlighted left inferior frontal and posterior temporal regions—approximately where the classical model would have predicted Broca’s and Wernicke’s areas. The result was useful, especially for presurgical mapping. It did not show that English lived in two illuminated blobs. The contrast isolated processes that differed between comprehended and uncomprehended speech while subtracting much of the auditory input shared by both.
Every activation map contains that logic. A region that appears in a contrast is not necessarily exclusive to the named function, and a region that disappears may be active in both conditions. Localization remains essential, particularly in clinical work, but localization answers where a manipulation changes activity. It does not by itself answer what computation the tissue performs or how the system produces language.
Lesion evidence has the complementary limitation. A damaged node can disable a network; a severed pathway can make intact nodes useless to one another; diaschisis can reduce function in distant tissue; and vascular lesions remove several structures at once. Functional imaging, stimulation, diffusion imaging, electrophysiology, and careful behavioral analysis each constrain the others. No single method gets to declare the module.
The emerging picture is not that language is everywhere. It is that language is produced by a structured network with reproducible hubs, pathways, and asymmetries. The network is distributed without being diffuse.
49.14 A language system can move
The devastating effects of an adult left-hemisphere stroke can make language lateralization look inevitable. If language is a human universal and both hemispheres begin with similar cortical tissue, why does the right hemisphere not simply take over?
Development provides the answer, but not a simple one.
Children who experience a major left-hemisphere stroke before or around birth often develop language far more successfully than adults with lesions of comparable size. Functional imaging shows that sentence processing can become organized in right-hemisphere regions homologous to the usual left frontal and temporal language network [@newport2022]. In rare cases of severe epilepsy, a diseased hemisphere is functionally disconnected or removed during childhood. When the left hemisphere is affected early enough, the remaining right hemisphere can support speech and language that are strikingly effective in daily life.
This does not mean the hemispheres are interchangeable spare parts. Outcomes vary. Subtle weaknesses may appear in complex syntax, rapid lexical retrieval, verbal working memory, discourse, or other demanding tasks. Seizure burden, the health of the remaining hemisphere, age at injury, the timing of surgery, and the child’s opportunities for communication all influence development. A brain that has endured years of uncontrolled epilepsy differs from a healthy brain experimentally reduced to one hemisphere.
Nor does plasticity mean that language tissue moves after it has been destroyed. The infarct remains. What changes is the developmental allocation of function in surviving circuits. An infant’s network has not yet completed the long process of specialization that makes the adult system efficient and constrained. Inputs that would normally strengthen left-hemisphere language pathways can instead strengthen right-hemisphere homologues. The new organization is built through use.
This helps explain why timing matters. In the adult, language is supported by highly practiced representations and long-range connections that have been shaped over decades. Moving the function would require recreating not only local computations but the network of relationships among them. Surviving regions can compensate, strategies can change, and perilesional tissue can recover, but plasticity must work within an already committed architecture.
The right hemisphere nevertheless contributes to adult recovery. Early after stroke, activation may increase in right homologues and in spared left-hemisphere regions. Whether right-hemisphere recruitment is helpful depends on the lesion and stage of recovery; it may provide useful support in some patients and reflect inefficient compensation in others. Recovery is not a contest in which the “good” left hemisphere must suppress the “bad” right one. It is a network trying to restore useful communication with the tissue that remains.
The practical lesson is humane as well as scientific. A first examination after stroke is not a final verdict. Aphasia can change substantially over weeks, months, and years. Therapy supplies repeated, structured opportunities for surviving circuits to solve communicative problems. Conversation partners also become part of the intervention by slowing down, confirming meaning, accepting gesture or writing, and helping the patient use preserved routes. Once again, the relevant control system exceeds the individual brain.
The ability of a young right hemisphere to support language is sometimes taken to mean that cortical regions have no intrinsic specializations. The severe aphasia following adult left-hemisphere damage is then taken to mean the opposite—that language was rigidly localized from the beginning.
Both conclusions are too strong.
Cortical areas enter development with differences in connectivity, cytoarchitecture, sensory input, gene expression, and timing. These biases make some developmental outcomes more likely than others. Experience then amplifies, redirects, or occasionally overrides those biases. Early injury can change which circuits receive the relevant input and practice; late injury occurs after a network has become deeply specialized.
Plasticity and specialization are therefore not opposites. Plasticity is one of the processes by which specialization develops.
49.15 Language as closed-loop control
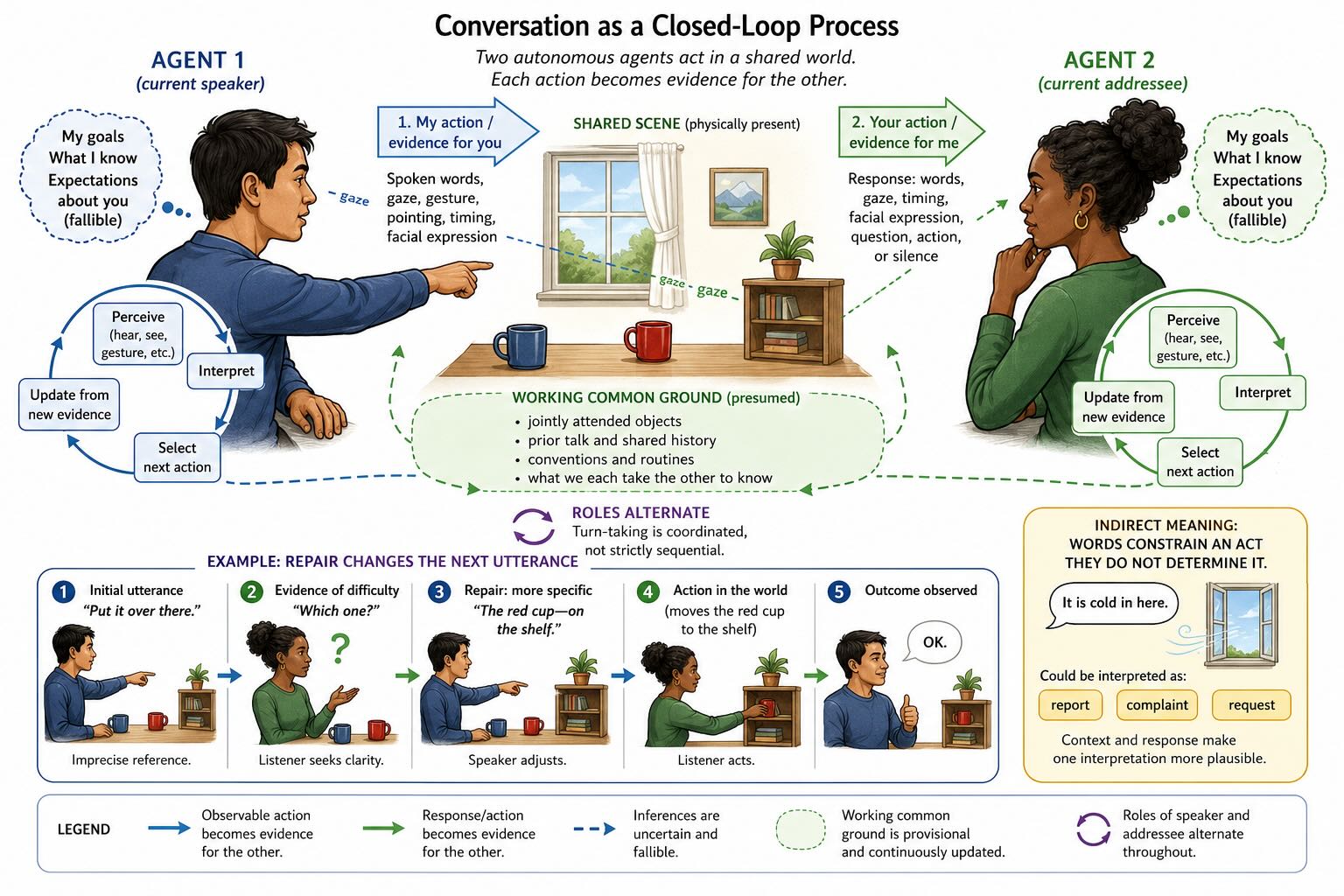
So far the chapter has sometimes described a signal moving from speaker to listener. Real communication is rarely that linear. It is a coupled feedback process.
A speaker acts with an intended effect: warn this person, identify that object, obtain help, establish agreement, make the listener laugh, or change what the listener expects. The utterance is selected partly from an estimate of what the listener can see and already knows. The listener responds through gaze, timing, facial expression, questions, action, or silence. Those responses become new sensory evidence for the speaker, who continues, stops, repeats, elaborates, or repairs.
The same expression can therefore be adequate for one listener and useless for another. Put it over there works when speaker and listener share attention to the same objects. It fails on a telephone. She did it again depends on a history both parties can retrieve. Communication succeeds not because words contain a complete message but because words constrain an interpretation within a jointly structured situation.
This is control in the limited sense defined at the beginning of the chapter. The speaker acts to alter a variable in the listener, observes imperfect feedback, and adjusts. The listener is not a passive target. They test the utterance against their own goals, knowledge, and trust in the speaker, and their response alters the speaker in return. Conversation is therefore better represented as two autonomous systems coupled through sound, gesture, gaze, and action than as a package sent through a conduit.
The coupling need not require an explicit inner theory of the other person at every moment. Habit, convention, shared routines, and immediate feedback solve much of ordinary conversation. Yet flexible language repeatedly exceeds those shortcuts. A speaker may need to estimate which referent this listener will choose, whether an indirect request will be recognized, what evidence the listener lacks, or how the utterance will be interpreted by someone with different loyalties. A listener asks the complementary question: why did this person say that, here, to me?
That structure places limits on the metaphor of language as a code. Language certainly uses conventional mappings, and code-like descriptions are useful for phonemes, words, and some grammatical relations. Ordinary utterances, however, remain underdetermined. Speakers omit what context supplies. Irony can communicate the opposite of a literal sentence. A question can function as a request. It is cold in here can be a report, a complaint, or an instruction to close the window. The acoustic or visual signal constrains meaning; it rarely specifies the intended act by itself.
Language is thus joint action carried by learned conventions. Its success depends on physically instantiated estimates of another agent, not on a detached symbolic copy of that agent hidden in one cortical box. This is why Chapter 50 follows. To communicate flexibly, a brain must sometimes estimate what another agent perceives, knows, expects, or falsely believes—and recognize that the other agent is estimating it in return.
49.16 What language did to the niche
Once a community possessed a sufficiently expressive language, the selection pressures surrounding its members changed.
A novice no longer had to learn only by trial, observation, and imitation. An expert could direct attention to the relevant feature, name a category that crosses superficially different examples, explain a sequence that could not be inferred from its visible result, and warn about a mistake before the novice made it. A group could plan an action whose parts were distributed among people and delayed in time. Norms could be stated, violations reported, alliances negotiated, and obligations remembered collectively.
Language also increased the value of knowledge that was difficult to acquire individually. A rare observation could spread without everyone repeating the dangerous encounter. Information about a distant water source, a poisonous plant, an animal’s seasonal movement, or a person’s unreliability could travel through the network. The discovery outlived the discoverer if others continued to tell it.
This did not make speech automatically truthful. A system that can report unseen events can also lie about them. A reputation network creates incentives to manipulate reputation. Instructions can coordinate care or violence. Myths can preserve useful norms or stabilize false beliefs. Language increased the power of one brain to alter another; it did not guarantee that the alteration served the listener.
That ambiguity is evolutionarily important. Listeners cannot simply absorb every utterance. They evaluate speakers, compare reports, notice inconsistencies, demand evidence, and track who has been reliable. Speakers, in turn, manage impressions and tailor claims to audiences. The social niche becomes partly an epistemic niche: survival depends not only on obtaining information but on deciding whose information to trust.
Language also became a new developmental demand. A child born into a linguistic group had to learn thousands of conventional forms, infer grammatical patterns, master turn-taking, distinguish literal from indirect meaning, and discover which ways of speaking marked age, status, intimacy, or group membership. Individuals better able to learn and use the system would gain access to more of the network’s knowledge and cooperation. The cultural invention therefore created selection pressures on attention, memory, vocal control, social motivation, and learning.
The arrows run in both directions. Biological changes made richer communication possible. Richer communication altered the cultural niche. The altered niche increased the value of biological capacities that supported language. Meanwhile, languages themselves changed as they passed through generations of learners.
Animals create niches that modify animals. Speaking animals create niches made partly of words.
49.17 Coda: the network speaks
We began with a forager returning from beyond the ridge. Her achievement is easy to overlook because human language feels effortless. The speaker moves air. The listener hears a sentence. A plan changes.
But the event crosses almost every level developed in this book. Metabolic systems support the brain and respiratory muscles. Auditory pathways transform pressure waves into neural activity. Motor systems control breath, larynx, tongue, and face. Basal ganglia select and sequence; cerebellum predicts and corrects. Temporal cortex analyzes learned sound patterns. Frontal and parietal networks maintain, combine, and route them. Memory supplies concepts and episodes. Social systems estimate what another person knows. A culturally inherited language provides the conventional forms. Two brains enter a feedback loop, and bodies act differently because of it.
The old localization story captures only a thin slice of that event. Broca’s and Wernicke’s discoveries remain foundational because lesions revealed that different parts of the left hemisphere make different contributions. Stroke syndromes remain indispensable because they show what a successful system normally hides. Functional imaging and stimulation make those dependencies visible in living brains. Yet language does not sit in Broca’s area, pass through a wire, and arrive in Wernicke’s area. It is assembled by a lateralized, distributed, plastic network that reaches from brainstem and basal ganglia through sensory, motor, temporal, parietal, and frontal cortex.
The hemispheres are not equivalent, but neither are they rival personalities. The most consequential asymmetry concerns the left hemisphere’s strong role in speech production, phonological sequencing, and linguistic combination. The right hemisphere contributes prosody, context, discourse, and social interpretation. The corpus callosum ordinarily makes the division invisible by allowing the partners to exchange information. Split-brain experiments are interesting because they interrupt that exchange, not because they reveal two complete people waiting inside every skull.
Evolution did not begin with the cortical map. It began with animals already communicating through bodies and voices, already sequencing actions, already tracking one another, already learning from social partners. In the hominin lineage, improved voluntary vocal control, gesture, shared attention, cooperative dependence, and cultural transmission formed a positive feedback loop. No single hypothesis explains the whole transition because no single trait became language. Old systems were coupled into a new one, and the new cultural system then helped reshape the brains that learned it.
Language is often described as a faculty possessed by an individual. In one sense it is: your brain contains knowledge of words, constructions, and conversational practices. In the more important sense, however, a language exists between individuals. A private word that no one else understands is not yet a useful word. Grammar is stabilized across a community. Meaning depends on shared histories and coordinated expectations. Every speaker inherits most of the system from people who inherited it from others.
The network remembers. Language lets the network speak.
This is why the human difference cannot be located solely in a larger brain or a special patch of cortex. Human language turns other brains into part of the action loop. It allows one nervous system to warn, instruct, promise, recruit, deceive, comfort, and teach another. It makes knowledge portable across bodies and persistent across generations. It is one of the principal mechanisms by which a murder of brains becomes more capable than any brain in it.
The next chapter takes one further step. Language works because speakers and listeners treat one another as agents with perspectives, knowledge, goals, and intentions. The most important object in the human niche is another control system. To use words effectively, a brain must predict that system—and, eventually, recognize itself as one.
This chapter has moved from animal communication through language evolution, hemispheric specialization, stroke, and cultural inheritance. The evidential foundations are not equally firm.
We are confident that: human language is biologically enabled but culturally acquired; speech and language are not the same thing; signed languages recruit a strongly left-lateralized language network; most people show a left-hemisphere bias for speech production and phonological processing; language nevertheless depends on bilateral and subcortical systems; left middle cerebral artery strokes produce recognizable but variable aphasic syndromes; the persistent deficits called Broca’s and Wernicke’s aphasia usually reflect damage to networks larger than the textbook areas; reading recruits and reorganizes evolutionarily older visual and language circuits; and early injury can shift language development toward the right hemisphere.
We have good reason to think that: language evolved through the integration of several preexisting capacities rather than through one new module; gesture and multimodal communication played an important role; increasing voluntary vocal control was crucial to speech; shared intentionality and cooperative dependence increased the value of informative communication; cultural transmission helped regularize and structure languages; lateralization improves the efficiency of densely interacting computations; and language changed the developmental and selective niche of later humans.
We remain genuinely unsure about: when anything recognizably like modern language first appeared; whether gesture, song-like vocalization, or some inseparable multimodal system carried the earliest conventional symbols; the relative contributions of cooperative breeding, tool teaching, pair bonding, group coordination, gossip, and ritual; which aspects of grammatical structure require language-specific biological biases; why language lateralization settles on the left in most individuals but not all; how closely Neanderthal communication resembled ours; and how the many components crossed the threshold into the open-ended system humans now inherit.
The uncertainty is not a reason to retreat to the old map of two language centers. It is a reason to keep the levels connected. Genes build developing circuits. Bodies act and signal. Brains specialize through experience. Communities stabilize conventions. Languages change as they are learned. Each level constrains the others, and none is the whole explanation.