16 Signaling
Overview
The previous units have established that the brain is an embodied control system that regulates the body to maintain homeostasis and to facilitate allostasis. In the sections on the circumventricular organs, the nucleus of the solitary tract and, in particular, the hypothalamus, we watched that control system reach out into the body and the body signal back: circulating molecules in the blood reporting the state of the periphery to the brain, and hormones released by the brain acting on distant tissues — and, returning, acting on neurons themselves. This unit is about the mechanisms underneath all of that. The word for the whole business is signaling, and the argument of the unit is contained in a single observation: signaling happens across an enormous range of timescales, from thousandths of a second to days, and the neuron is a participant at every scale — but for different reasons at each.
That observation is not original to me. The neuroscientist Robert Sapolsky has made a habit of asking, about any piece of behavior, on what timescale do its causes operate? — what happened a second before, an hour before, a lifetime before, an evolutionary epoch before. The same discipline applies inside the single neuron. A neuron responds to a glutamate molecule in a millisecond and to a steroid hormone over hours, and these are not the same kind of events. They are different mechanisms, serving different purposes, operating at different speeds, and a neuron evolved to respond to all of them. Most introductions to neural signaling collapse this range down to its fastest layer — the firing cell, the spike or action potential, the synapse — and treat everything slower as background. We will, of course, get to fast signaling later in this unit. But I want to resist the implicit notion that all signaling is just spikes from the start, because the fast electrical layer is not the foundation of signaling. It is the most recent, most specialized, and by a wide margin most expensive part of a much older system.
16.1 Why signal at all
Begin with the reason a cell would need to signal another at all. The first unit’s answer was movement: a nervous system exists to coordinate the activity of a body so that the organism can move adaptively — toward food, away from threat, toward a mate. Even the simplest coordinated movement requires that one part of the system communicate with another. A circuit that alternates a limb forward and back must have some cells exciting the muscles that flex and others inhibiting them, in turn, in time. Coordination is communication, and communication of exactly two basic messages — excite and inhibit — is the raw material from which all of it is built.
This is not as impoverished as it sounds. Two opposed messages, delivered together, do not cancel into nothing: they sum, and a target cell driven by a mixture of excitation and inhibition can be pushed anywhere along a continuous range, held just below threshold, or made to fire at any rate in between. A limb does not simply flex or extend; it moves smoothly, with graded force, because the balance of push and pull is continuously adjusted. The vocabulary is binary; the messages it composes are not. Everything in this unit is ultimately machinery for sending those two messages, in the right place, at the right time, in the right proportion, and on the right timescale.
16.2 Forms of signaling
Cells signal one another in several physically distinct ways. The differences come down to how far the message travels and how it gets there, and the four basic forms run from the most far-reaching to the most intimate:
- Endocrine signaling sends a molecule — a hormone — into the bloodstream, to be carried throughout the body and act on any cell bearing the right receptor. It is slow, because it depends on circulation, and broad, because the blood goes everywhere.
- Paracrine signaling is local: a molecule is released and diffuses a short distance to neighboring cells. Synaptic transmission, as we will see, is essentially a tightly controlled form of paracrine signaling.
- Autocrine signaling is a cell signaling itself — its own released molecule binding receptors on the very cell that released it.
- Direct signaling dispenses with the released molecule altogether. Through gap junctions, the interiors of two adjacent cells are physically continuous, and ions or small molecules pass straight from one to the other. This is the basis of the electrical synapse, the fastest form of communication between cells.
A point that will recur throughout the unit is already visible in these categories. The systems of the body are named for their messengers — hormones in the endocrine system, cytokines in the immune system, neurotransmitters and neuromodulators in the nervous system — but the messengers do not respect the boundaries. Many of the very same molecules serve in more than one system. This is the first hint of a principle we will develop carefully: what a signaling molecule means is not a fixed property of the molecule. We have, in fact, already seen this principle at work. In the previous unit, circulating hormones reported the body’s state to the brain and acted directly on neurons. The molecules that did so were not built for the brain; they were endocrine signals that neurons happened to be equipped to read.
16.3 Where the vocabulary came from
Before going further into the machinery, it is worth a brief digression on how old this vocabulary is — because it is far older than the nervous system itself.
Consider the marine sponge. It is about as simple as an animal gets: no neurons, no nervous system, no electrical transmission of the kind we are about to study. It is a filter feeder, drawing water through its channels to strain out bacteria. Yet it faces a control problem, because those channels clog, and when they do the animal clears them with a slow whole-body contraction that looks, reasonably enough, like a sneeze. The sneeze must be coordinated, and the sponge has no nerves to coordinate it. What it has is chemistry. The contraction depends on the release of glutamate, and it can be suppressed by GABA — the very molecules that serve as the principal excitatory and inhibitory transmitters of your own brain. The animal with no nervous system is already running its coordination on the chemical signals you are using to read this sentence.
The molecules of signaling, in other words, are inherited, not invented. The nervous system did not design its vocabulary from scratch; it specialized in the use of molecules that cells had been signaling with for a very long time. And we can say something about where those molecules came from. Every cell is a web of chemical reactions, and the concentration of any reaction’s product is itself a readout of how fast that reaction has been running. A molecule sitting at some concentration is therefore already carrying information about the state of the machinery that makes it — already, in a minimal sense, a signal. Evolution did not have to invent messengers; it had only to repurpose molecules that were already reporting on the cell’s internal state.
16.4 Membranes and the cost of a mobile cell
That is enough about the molecules. The deeper question for this unit is not where the signals came from but why signaling has the character it does — and in particular why its fastest layer is so costly. To answer that we have to start not with neurons but with the boundary that makes a cell a cell: the membrane.
Without a membrane there is no cell — nothing to distinguish a single-celled organism like an amoeba from the pond water around it. The membrane is the partition that separates life from its environment. But the moment a cell wraps itself in a membrane, it has a problem, because a perfectly sealed membrane would be a tomb. A cell cut off from its surroundings could not respire, take in nutrients, or expel waste. A membrane therefore cannot be a perfect barrier; it must be permeable, punctuated by pores and channels and receptors that allow the regulated traffic of matter and information. Permeability is not optional. It is forced on the cell by the need to stay alive.
And permeability immediately creates a vulnerability. A cell is full of dissolved solutes, so its interior is more concentrated than fresh water. Put such a cell in pond water and the concentration gradient will drive water in across the permeable membrane, swelling the cell and threatening to burst it. (This is why dried beans, soaked overnight, come out swollen the next morning.) This osmotic threat is a genuine engineering crisis, and evolution solved it in two very different ways. Plants solved it structurally, encasing each cell in a thick rigid wall that resists the pressure. The solution works, but it has a cost: a cell in a rigid box cannot move. Plants are sessile, and must make their own food where they stand because they cannot go and find it.
Animals took the other path. An animal moves — to forage, to escape, to find a mate — and movement is incompatible with a rigid wall. So the animal cell keeps a thin, flexible membrane, and pays for that flexibility by managing the osmotic threat actively rather than structurally. It continuously pumps ions across its membrane, manipulating the electrochemical gradients across the boundary to keep its interior from flooding. Pumping ions against their gradients is uphill work — like pumping water up into a water tower against gravity — and uphill work costs energy. The animal cell, having given up the wall for the freedom to move, must spend metabolic energy continuously, for its whole life, simply to hold its own contents in balance.
Here is the connection that matters for this unit. Those pumped ion gradients — a great deal of sodium held outside the cell, a great deal of potassium held in — are a store of potential energy maintained across the membrane. The neuron’s fast electrical signals are, mechanically, the controlled discharge of that store: a channel opens, ions rush down the gradient, and the membrane voltage changes. The machinery that an animal cell evolved to survive without a wall is the same machinery the neuron would later exploit to signal quickly. I do not mean that the action potential is osmotic regulation, or that managing osmolarity is the origin of neural communication — that would be an overstatement of a real continuity. The truth is more modest and more interesting: fast electrical signaling did not require a new energy source. It spends a store of energy that animal cells were already maintaining for an older reason, and it inherits that store’s continuous metabolic price.
That price is substantial. As we have learned, the human brain consumes roughly twenty percent of the body’s energy at rest. A large share of that goes to a single molecular machine — the sodium–potassium pump — doing nothing more glamorous than maintaining the disequilibrium that keeps neurons poised to fire. We asked, at the end of the first unit, what it costs to buy prediction. Here is part of the answer, in metabolism: the fast end of signaling is fast precisely because it is expensive, and the brain must cover that expense for as long as it is alive.
16.5 How a signaling molecule reaches a neuron
When a signaling molecule reaches a neuron, it can act by entering the cell or by binding to a protein in the membrane. These routes differ markedly in their mechanisms and speed.
Small uncharged or fat-soluble molecules can diffuse through the lipid bilayer. Steroid hormones—including cortisol, estrogen, and testosterone—are important examples. Some other molecules enter with the help of dedicated transporters, whereas molecules that remain outside the cell act through membrane channels or receptors.
Cortisol shows how the same signaling molecule can act through more than one route. Because cortisol is fat-soluble, it can cross the neuronal membrane and bind a glucocorticoid receptor in the cytosol. The cortisol–receptor complex then enters the nucleus and alters gene transcription, producing effects that develop over hours and may persist much longer. Cortisol can also act through receptors associated with the cell membrane, triggering intracellular signaling within seconds to minutes. In the example shown in Figure 16.1, this faster pathway stimulates production of the endocannabinoid 2-AG, which travels backward across the synapse and suppresses neurotransmitter release from the presynaptic terminal.
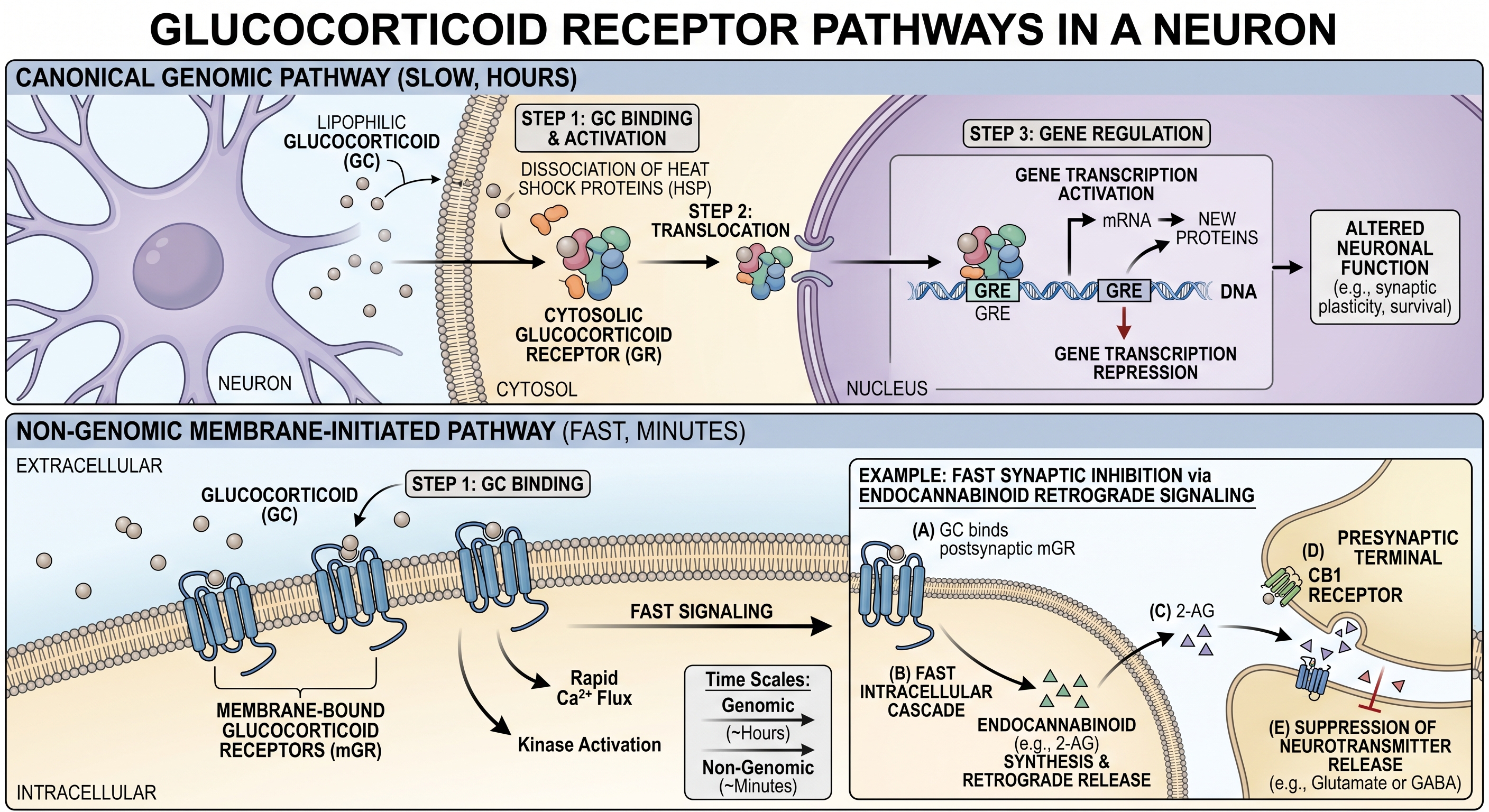
The membrane-initiated pathway illustrates the general logic of metabotropic signaling: binding at the cell surface activates intracellular intermediaries, which may regulate enzymes, ion channels, or other cellular processes. An ionotropic receptor is more direct—the receptor is itself an ion channel, and binding opens the channel. Leak channels require no activating signal and remain open, providing baseline permeability to selected ions. These distinctions are developed in detail, together with voltage-gated channels, in Section 18.5.
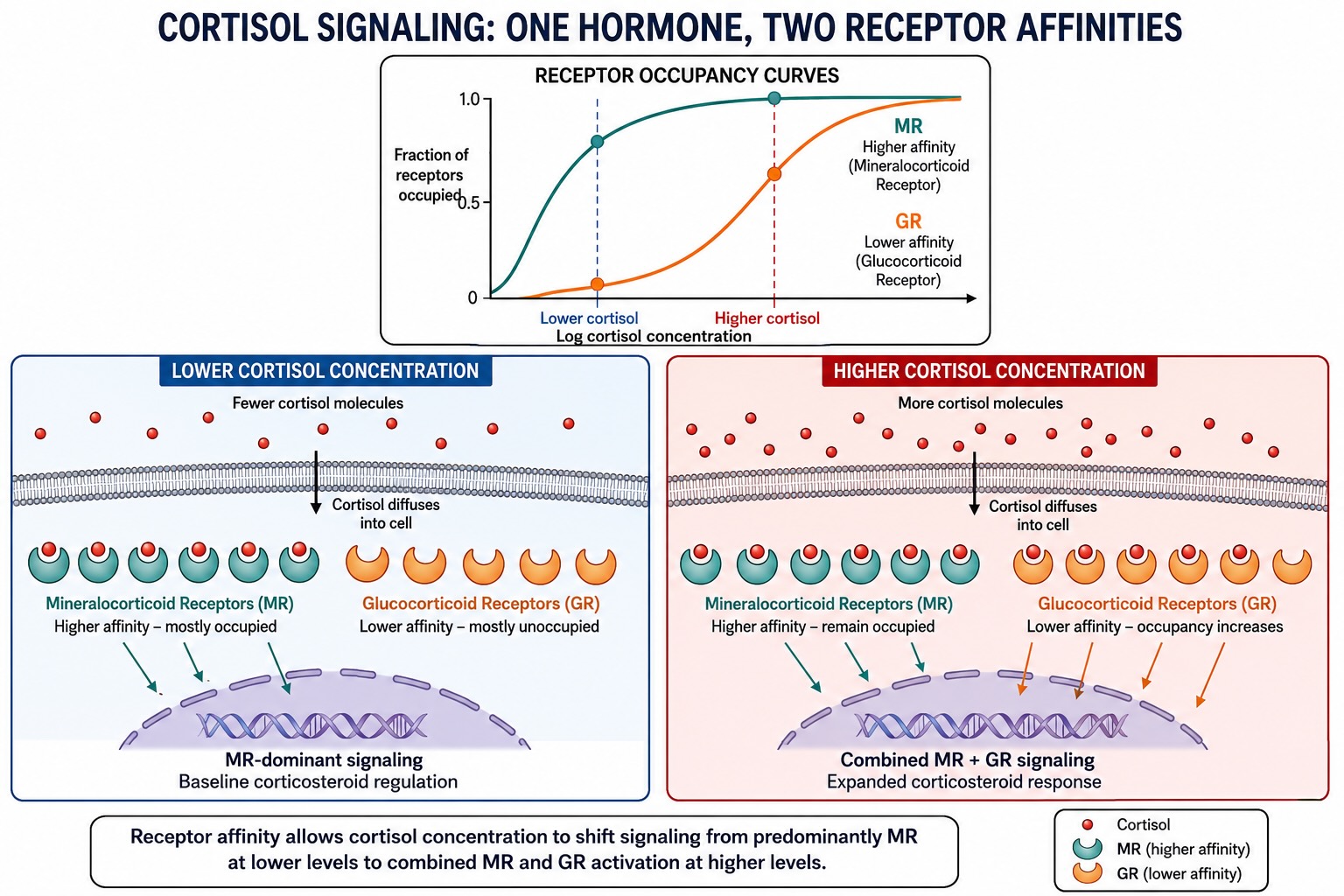
A signaling molecule therefore does not carry a single meaning. Its effect depends on the receptor and cellular machinery it encounters. Moreover, when several receptors recognize the same molecule, they may not bind it equally strongly. That binding strength—the receptor’s affinity—helps determine which pathways are recruited at a given concentration. Thus, the concentration of the signaling molecule itself becomes a message. A given molecule is often read by several receptor types that differ in their affinity — in how tightly they bind it, and therefore in the concentration at which they respond. A high-affinity receptor is occupied even when the molecule is scarce; it is sensitive to the molecule’s baseline, tonic level. A low-affinity receptor stays empty until the molecule becomes abundant; it responds only to a surge, a phasic burst. Run a single molecule past both, and the cell can tell the difference between “a little, all the time” and “a lot, just now” — because the two conditions engage different receptors and so produce different effects. Cortisol again provides the clean example: its high-affinity mineralocorticoid receptors are largely occupied at the body’s resting cortisol level and track normal daily regulation, while its lower-affinity glucocorticoid receptors are recruited only when cortisol spikes under stress, switching on a distinct genetic program. The hormone says one thing at rest and another under stress, using the same molecule, by means of receptors tuned to different concentrations. Affinity makes concentration an information channel in its own right.
16.6 The argument and the unit ahead
Step back and the shape of the argument is the timescale itself. An electrical synapse passes a signal between coupled cells almost instantaneously. A glutamate molecule at an ionotropic receptor changes a neuron’s voltage in a millisecond or two. A neuromodulator working through a surface receptor and an internal cascade shifts the neuron’s excitability over seconds. A steroid hormone reaching the genome remakes the neuron over hours and leaves a trace for days. A hormone circulating in the blood sets the body’s operating state over still longer spans. These are not five versions of one thing. They are genuinely different mechanisms, and the neuron takes part in all of them — using the fast ones to carry information moment to moment, and the slow ones to regulate how that information is carried and whether it leaves a lasting mark. This is the sense in which, following Sapolsky, we will examine the neuron’s response along many timescales at once.
The chapters ahead descend through these timescales in turn. We begin with the cellular cast — neurons, glia, and the synapse between them — because we should know the players before we watch them act. We then build the fast electrical layer from first principles: membranes and gradients in detail, the channels that open and close, the way a neuron sums its inputs, and the action potential that carries the result down the axon — along with the metabolic price we have just previewed. From there we widen into the slower chemical environment in which all that fast signaling is bathed: neuromodulation, the diversity of receptors, and the difference between a signal aimed at a single synapse and one released to drift across many cells at once. We give the glia their own chapter, and with them the broader traffic between the nervous system and the body’s other signaling systems, because the synapse is not the only place signaling happens. And we end where signaling becomes memory — the activity-dependent changes that let a synapse record its own history, how those changes are reinforced, and a first look forward at how a brain might learn to assign credit for a reward that arrives only after the act that earned it.
Throughout, the molecule remains as it was in the sponge. What grows — what made a nervous system possible, and what organizes everything ahead — is the range of timescales over which a neuron can be made to respond, and the receptors that decide, in each case, what a simple chemical signal is going to mean.