23 Pain and Temperature
Nociception, Valuation, and Control
Pain begins within somatosensation but does not end there. Its peripheral receptors lie in the skin, muscles, joints, and viscera; its first-order neurons enter through dorsal-root or cranial-nerve pathways; and much of its ascending traffic uses the anterolateral system introduced in the preceding chapter. Yet the resulting experience includes more than the location and intensity of a stimulus. Pain is unpleasant, commands attention, alters autonomic state, and organizes protection, escape, guarding, and learning.
Temperature shares much of this peripheral and ascending machinery, but ordinary warmth and cooling are not simply weak forms of pain. They report conditions at the body surface, guide contact with objects and environments, and help organize behavior before tissue is threatened. Only as heat or cold moves outside a tolerable range does thermal sensation become strongly coupled to nociception and pain.
Pain makes particularly clear a principle that applies throughout the sensory systems. Sensing is not a neutral recording stage followed later by valuation and action. Sensory pathways are organized around what information means for the animal. Pain makes that organization unusually explicit because information about tissue threat arrives already coupled to urgency and aversion.
The chapter therefore begins with ordinary temperature sensation and then turns to nociceptors and the fast and slow afferents that carry their signals. It returns to the anterolateral system developed in the preceding chapter, follows its signals into several interacting control networks, and then closes the loop through the brainstem and spinal circuits that regulate the next incoming signal. The final sections consider what happens when pain persists or becomes partly uncoupled from immediate tissue damage.
The International Association for the Study of Pain defines pain as an unpleasant sensory and emotional experience associated with, or resembling that associated with, actual or potential tissue damage [@raja2020revised]. The wording matters. Pain is an experience, not a direct readout of an injured tissue.
Nociception is the neural encoding of noxious stimuli—events that actually or potentially damage tissue. It includes transduction at nociceptors, activity in peripheral afferents, spinal and brainstem processing, and ascending traffic. Nociceptive activity can occur without pain, as when noxious stimulation evokes neural or reflex responses during adequate general anesthesia. Pain may also persist when no current tissue lesion or external noxious stimulus fully accounts for it, as in some phantom-limb and chronic pain states.
The distinction does not imply that pain is unreal when no injury can be found. It means that activity in a nociceptor is neither necessary nor sufficient, by itself, to explain the experience. Pain must be understood at the level of the whole organism, including its sensory pathways, bodily state, learning history, context, and available actions.
23.1 Temperature before tissue threat
Thermal sensation begins largely in free nerve endings in the skin. Across an ordinary range, warming and cooling can be clearly perceived without signaling tissue damage. These afferents report both the direction of change and the temperature reached relative to the skin’s recent adaptation. A metal railing on a cold morning and a warm cup in the hand therefore provide information about the environment at the point where it meets the body, even when neither contact is painful.
The molecular front end was introduced in the preceding chapter. Members of the TRP channel family respond over partly overlapping thermal ranges: TRPM8 contributes to cooling and menthol sensation, while TRPV1 responds to noxious heat, protons, and capsaicin [@caterina1997trpv1; @mckemy2002trpm8]. These examples should not be turned into a one-channel-per-temperature code. Populations of thermoreceptive and polymodal afferents together represent warming, cooling, and increasingly threatening thermal conditions.
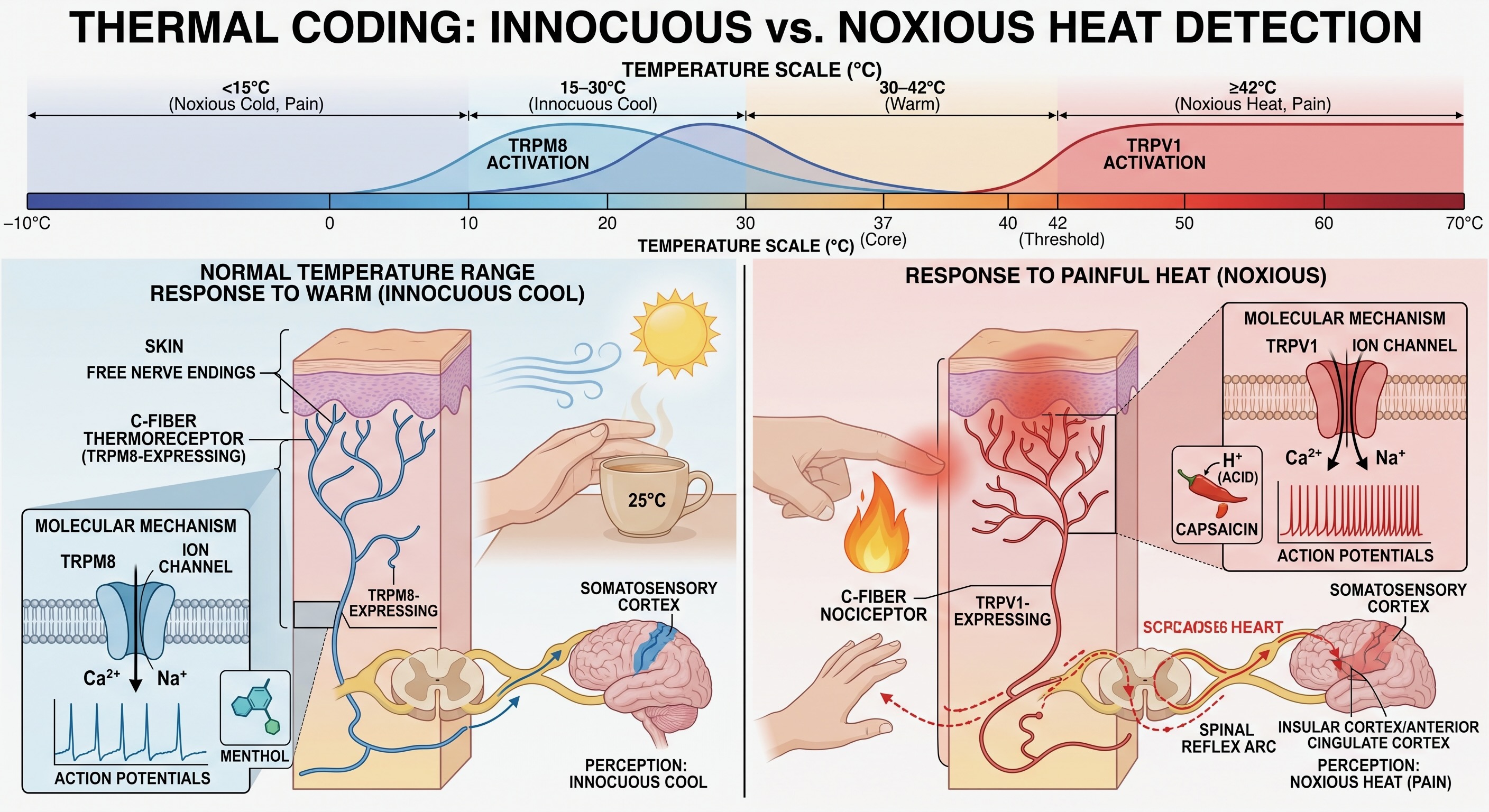
Cutaneous temperature also exposes the seam between exteroception and interoception. At the skin, temperature helps identify the thermal properties of an object or environment and guides contact, withdrawal, clothing, shelter seeking, and other actions. Core temperature, by contrast, is an internally regulated variable whose homeostatic control was developed in Unit II. The physical variable is the same, but a receptor at the body surface and a sensor contributing to core-temperature regulation enter different control loops.
As heat or cold moves farther outside the safe range, high-threshold thermal afferents and polymodal nociceptors are increasingly recruited. The transition is not a single fixed temperature at which an innocuous receptor hands the signal to a pain receptor. Adaptation, tissue condition, inflammation, stimulus duration, and the populations of afferents recruited all matter. Innocuous and noxious thermal signals nevertheless share much of their spinal entry and ascend mainly through the anterolateral system, which is why temperature and pain are clinically lost together after some spinal lesions.
23.2 Nociceptors and tissue threat
Many nociceptors end as free nerve endings: branching terminal axons without an encapsulating corpuscle. They are simple in gross morphology but not in molecular specialization. Different endings respond preferentially to high-threshold mechanical deformation, noxious heat or cold, chemical irritants, or combinations of these conditions.
Some damaging stimuli act directly on transduction channels. Noxious heat, protons, and capsaicin can open TRPV1, for example. Tissue injury also changes the chemical environment around an ending. ATP, protons, bradykinin, prostaglandins, histamine, cytokines, and other mediators released by damaged cells, blood vessels, and immune cells can activate nociceptors or increase their responsiveness [@basbaum2009cellular]. The familiar tenderness around a cut or burn is therefore not simply a more intense version of the original stimulus. The tissue has changed the gain of its own warning system.
This increase in peripheral responsiveness is peripheral sensitization. A nociceptor may begin firing to weaker stimulation, respond more strongly to the same stimulation, or remain active in the inflammatory environment after the original insult. The result is protective: movement and contact around an injured site become aversive, encouraging the animal to guard the tissue while it heals. The same mechanism can also contribute to persistent pain when inflammation or peripheral nerve activity continues.
The term nociceptor is preferable to pain receptor. These endings detect conditions associated with tissue threat. Whether their activity becomes pain depends on what happens in spinal, brainstem, thalamic, and cortical networks.
23.3 Fast and slow pain
The preceding chapter placed nociceptive afferents within the broader Aα–Aβ–Aδ–C hierarchy. Proprioception and discriminative touch use the fastest lanes; nociceptive traffic travels mainly on the smaller Aδ and C fibers. A brief noxious stimulus to the skin can therefore produce two successive sensations carried predominantly by different peripheral afferents.
- First pain is sharp and relatively well localized. It travels mainly on thinly myelinated Aδ fibers.
- Second pain is slower, burning, aching, or throbbing, and often less precisely localized. It travels mainly on unmyelinated C fibers.
Stubbing your toe can reveal the sequence: an immediate sharp sensation, a short interval, and then a slower ache that lasts. The gap reflects the different conduction velocities of the two axon populations. Myelinated Aδ activity reaches the spinal cord and brain before the slower C-fiber activity.
The difference in timing does not mean that the body postponed its first protective response until pain reached cortex. Withdrawal can begin through spinal circuits before conscious perception is complete. The two afferent populations are nevertheless suited to partly different roles. Rapid Aδ input can help identify and localize a threat, while sustained C-fiber input can prolong attention, guarding, and recuperative behavior.
These are tendencies, not two sealed channels. Natural injuries usually recruit mixtures of afferents, and ongoing inflammatory, visceral, and neuropathic pain rarely arrives as a neat pair of sensations. The first-pain/second-pain distinction is most useful for understanding the response to a brief, experimentally controlled cutaneous stimulus.
23.4 Returning to the anterolateral system
The preceding chapter established the anatomical bookkeeping that matters clinically. Small-diameter first-order afferents enter through a dorsal root, may travel briefly in the dorsolateral tract of Lissauer, and terminate especially in laminae I and II and, for some afferents, in deeper dorsal-horn laminae. Local interneurons strongly transform this input, while many neurons that send axons toward the brain lie in lamina I and the deeper dorsal horn. Their axons cross through the anterior white commissure and ascend contralaterally. The local entry, synapse, and crossing are shown in Figure 22.2; a simplified ascending route is shown in @Figure 22.4. A broader schematic is presented in Figure 23.2.

The anterolateral system is broader than the spinothalamic tract and is not a dedicated pain cable. It carries thermal and nociceptive signals, but also itch and some less discriminative forms of touch. Its ascending axons arise from spinal neurons whose responses have already been shaped by local excitation, inhibition, convergence, and descending control. The distinction between Aδ and C fibers should therefore not be converted into a diagram with two peripheral wires ending in two cortical destinations.
The face follows a parallel cranial route. Nociceptive and thermal afferents travel mainly through the trigeminal nerve, enter the brainstem, and synapse principally in the spinal trigeminal complex. Second-order axons cross and ascend through trigeminothalamic and related pathways toward the VPM and additional thalamic and brainstem targets. The body and face therefore enter through different peripheral structures, but both feed distributed systems for discrimination, bodily regulation, arousal, valuation, learning, and action.
23.4.1 One event, several control problems
The fan-out in Figure 23.2 should not be read as four independent kinds of pain. It shows several weighted entry routes into overlapping control networks. The familiar distinction between a sensory-discriminative dimension and an affective-motivational dimension remains useful, but it compresses a richer architecture into two labels. A noxious event creates several related problems at once: Where is the threat? What condition is the body in? How much priority should the event receive? What defensive state should it recruit, and what should be learned for the future?
Touching a hot pan makes the sequence concrete. Spinal circuitry can begin withdrawing the hand before the event is fully perceived. Somatosensory systems identify the fingers involved and help direct withdrawal and later guarding. Posterior insular-opercular systems represent the changing thermal and nociceptive condition of the body. Cingulate networks help interrupt the ongoing task and assign urgency to escape and protection. Parabrachial, amygdalar, hypothalamic, and PAG circuits recruit aversion, autonomic adjustment, defensive action, and learning. Descending projections then alter the transmission of subsequent nociceptive input, while withdrawal itself removes the hand from the heat. Neural feedback and action both close the loop.
No one branch contains the completed experience. The experience and the behavior it organizes depend on interactions among these systems, together with memory, context, attention, and the available actions. The branches differ in emphasis, timing, and connectivity, but they converge extensively.
23.4.2 Where is the threat? Somatosensory guidance
Lateral thalamic projections to primary and secondary somatosensory cortex contribute information about body location, timing, intensity, and stimulus properties. These representations matter because protective action must be spatially directed. The organism must orient toward the affected site, withdraw the appropriate body part, distinguish a focal external event from a diffuse internal disturbance, and later guard or inspect the tissue. The route is therefore better understood as discriminative-sensorimotor than as merely sensory.
Somatosensory cortex is not a cortical pain receptor. S1 and S2 respond to many innocuous events, and nociceptive information reaches them as part of a much broader somatic representation. In a magnetoencephalography study of brief laser stimuli, activity associated with first pain was especially prominent in S1, activity associated with second pain was especially prominent in anterior cingulate cortex, and S2 participated in both [@ploner2002cortical]. The result supports differentiated weighting within a distributed system; it does not show that Aδ fibers terminate only in S1 or that C fibers terminate only in cingulate cortex.
Hypnosis studies provide a related, but equally partial, dissociation. Suggestions that changed reported intensity were accompanied primarily by changes in S1, whereas suggestions that changed unpleasantness were accompanied primarily by changes in cingulate activity [@rainville1997pain; @hofbauer2001cortical]. The dissociation is asymmetric rather than clean: the intensity manipulation altered S1 but also produced changes elsewhere, and the null findings carry little weight given small samples and subtraction-based PET. Participants were also selected for high hypnotizability, and suggestion may in principle shift how pain is reported rather than how it is felt. Still, the results are useful because they show that components of pain can be reweighted. They do not divide pain into an objective signal in S1 and an emotional reaction somewhere else.
23.4.3 What condition is the body in? The operculo-insular route
The insula is a sheet of cortex buried deep within the lateral sulcus. In an intact lateral view it is covered by the frontal, parietal, and temporal opercula—literally, the lids over the insula. The cortical overview in Figure 21.3 opens those opercula to expose the insula and the neighboring secondary somatosensory region. This hidden geometry matters because the posterior insula and medial parietal operculum form a contiguous operculo-insular territory rather than two widely separated cortical islands.
The posterior and dorsal insula are especially important for thermal, nociceptive, and other somatic signals carried by the anterolateral system. Graded cooling produces temperature-related activity in the dorsal middle-to-posterior insula [@craig2000thermosensory]. Direct electrical stimulation of posterior insular sites can evoke nonpainful somatic sensations, warmth or cooling, and sometimes pain, with evidence of a rough body order [@ostrowsky2002insula; @stephani2011insula]. In a much larger series of cortical stimulations, painful experiences were rare overall but concentrated in the medial parietal operculum and adjacent posterior insula [@mazzola2012stimulation]. These findings provide stronger evidence for causal participation than activation alone, but they still do not identify a solitary pain center.
More anterior insular territories interact extensively with cingulate, prefrontal, autonomic, and limbic systems. Stimulation studies support a broad posterior-to-anterior shift: posterior sites more often yield somatic, thermal, or painful sensations, while more anterior and central sites are more likely to yield visceral or gustatory sensations [@stephani2011insula]. The gradient is useful, but it is not a rigid serial ladder in which posterior insula first records the body and anterior insula then manufactures a feeling. Responses overlap, many anterior sites are silent during direct stimulation, and pain depends on interactions extending far beyond the insula.
Human intracranial recordings make the parallel organization particularly clear. Thermonociceptive responses began at nearly the same time in posterior insula and amygdala, while responses in anterior insula began later and showed evidence of convergence from the two systems [@bastuji2018convergence]. The result does not prove that anterior insula creates pain, but it argues against a simple serial chain from sensation to emotion. Sensory and limbic routes are activated in parallel and interact as the event acquires meaning for the organism.
Pain belongs to somatosensation because it begins with signals from skin, muscle, joints, and viscera. It also overlaps deeply with interoception, because it reports the condition and vulnerability of the body itself. A painful event can alter heart rate, blood pressure, respiration, endocrine state, posture, facial expression, and the allocation of attention. These are not decorations added after a neutral sensation has been recognized. They are components of a coordinated protective response.
23.4.4 How much priority should the event receive? Cingulate cortex and action
The cingulate contribution is often described as the affective-motivational component of pain. The phrase is useful so long as affective does not mean an optional emotional coloring added to an otherwise complete sensation. Unpleasantness changes behavior. It interrupts the current task, creates a demand for relief, and biases the organism toward withdrawal, escape, guarding, help-seeking, or endurance when escape is impossible.
The broad term anterior cingulate cortex has been used inconsistently across the pain literature. Many nociceptive and action-related responses lie in territories now described as anterior or midcingulate cortex. Whatever the boundary chosen, the relevant regions interact with motor, autonomic, prefrontal, insular, and limbic systems. Their role is therefore better framed in terms of urgency, effort, anticipation, and action allocation than as a container for suffering.
Single-neuron recordings in macaques support that interpretation. Cingulate neurons can respond during anticipation of an avoidable noxious event and during performance of a pain-avoidance task [@koyama1998anticipation; @koyama2000avoidance]. The activity links nociception to what the animal is preparing to do. In rats, optogenetic inhibition of anterior-cingulate glutamatergic neurons abolished formalin-induced conditioned place aversion without altering nocifensive behavior during conditioning, although the effects of activating the same broad population differed between males and females [@jarrin2020differential]. These findings do not isolate a cingulate pain module; they show that nociceptive information enters systems that determine behavioral priority.
The hypnosis findings described above and historical experience with anterior cingulotomy are consistent with partial specialization, but neither supplies a clean localization experiment. Clinical series suggest that stereotactic lesions in the anterior cingulate region can reduce the burden of otherwise intractable pain, but the evidence is observational and heterogeneous [@sharim2016cingulotomy]. The procedure may alter attention, emotional reaction, action, and reported intensity in different proportions. The often-repeated claim that patients still feel exactly the same pain but simply cease to care about it is too tidy for the available evidence.
The cingulate branch therefore answers a control question rather than supplying a separate sensation: How strongly should this event interrupt current behavior, and what effort should be allocated to escape, protection, or relief?
23.4.5 What defensive state should be organized? Parabrachial nucleus, amygdala, hypothalamus, and PAG
The spinoparabrachial pathway supplies the clearest reason not to treat pain as a message delivered only to cortex. A major population of dorsal-horn projection neurons reaches the lateral parabrachial nucleus in the pons without going through a thalamic relay. The parabrachial complex also receives visceral and homeostatic information, placing nociceptive input in direct contact with systems concerned with the condition of the organism.
From the lateral parabrachial nucleus, signals reach the central amygdala, bed nucleus of the stria terminalis, hypothalamus, PAG, and other targets. Cell-type-specific work in mice shows that this fan-out can support different components of the response. Lateral parabrachial projections toward ventromedial hypothalamus or lateral PAG can promote escape, whereas projections toward central amygdala or BNST can support aversive learning [@chiang2020divergent]. These findings should not be mapped one-for-one onto the human brain, but they demonstrate the kind of biological organization that a single label such as emotion conceals.
The amygdala is not a fear center or a suffering center. It is a collection of nuclei in which nociceptive input interacts with context, prior experience, and other information about biological significance. In rats, activating the parabrachial-to-central-amygdala pathway can produce avoidance and other negatively valenced behavior without necessarily increasing simple nociceptive sensitivity [@cai2018brain]. In mice, inhibiting CeA-projecting parabrachial neurons reduced hypersensitivity after inflammatory or neuropathic injury without altering baseline nociception, whereas activating them produced hypersensitivity without a peripheral injury [@torresrodriguez2024pbncea]. Different central-amygdala cell populations can also increase or suppress pain-related behavior [@wilson2019dual]. The same gross structure can therefore participate in aversion, amplification, and suppression, depending on which cells and connections are engaged.
Hypothalamic outputs coordinate autonomic and endocrine adjustments, while the periaqueductal gray helps organize defensive modes and their motor expression. Central-amygdala neurons projecting to different PAG territories are physiologically diverse, and inflammatory pain alters the properties of some of these neurons in mice [@li2018ceapag]. Other rodent work shows that amygdala-to-PAG circuitry contributes to freezing and interacts with circuits supporting flight, while hypothalamic and parabrachial outputs recruit PAG and other brainstem effectors [@tovote2016midbrain; @chiang2020divergent]. PAG circuits also recruit descending systems that suppress or facilitate nociceptive transmission [@heinricher2009descending]. Defensive action and analgesia are not unrelated functions. An animal escaping immediate danger may temporarily reduce responsiveness to an injury, whereas a protected animal can afford to guard, rest, and attend to the damaged tissue.
The finest circuit dissection in this section comes predominantly from rodents. Comparative anatomy supports parallel spinothalamic and spinoparabrachial routes, while human intracranial physiology is consistent with rapid access to both posterior insular-opercular and amygdalar systems. The correspondence among species, cell types, and subnuclei is not exact. In human recordings, posterior insular and amygdalar nociceptive responses began with nearly identical latencies and converged later with anterior insular activity [@bastuji2018convergence]. The result supports parallel access to sensory and defensive-valuation systems without implying four sealed pathways or four independent experiences.
This branch answers a fourth control question: What coordinated defensive, autonomic, and learning state should this event recruit, and how should it change future behavior?
This account is independent of the contested claim that observing another person’s pain or experiencing social rejection reproduces first-person pain. Setting that controversy aside does not remove affect from pain itself. Aversion, urgency, autonomic adjustment, defensive action, and learning are part of the biological organization of first-person pain.
23.5 Closing the loop: pain changes the signal it receives
The ascending branches are not endpoints. The cortical, amygdalar, hypothalamic, and brainstem systems recruited by nociceptive input send signals back toward the spinal cord and alter the processing of the next input. Pain is therefore embedded in a recurrent control architecture rather than arranged as a one-way sequence from receptor to cortex.
Forebrain and hypothalamic systems recruit the periaqueductal gray (PAG) and related brainstem circuits. The PAG–rostral ventromedial medulla (RVM) pathway and descending noradrenergic pathways from the locus coeruleus reach the dorsal horn, where they alter the transmission of incoming nociceptive activity [@heinricher2009descending].
Descending control can inhibit nociceptive transmission. Endogenous opioid peptides act in the PAG, brainstem, spinal cord, and peripheral terminals. Within the dorsal horn, opioid receptors can reduce transmitter release from primary afferents and decrease the excitability of postsynaptic neurons. Morphine and related drugs exploit modulatory machinery that evolved for endogenous control.
The system is not exclusively analgesic. Descending pathways can also facilitate spinal nociceptive processing, increasing responsiveness in some contexts and pain states. Experiments in male mice have identified PAG-to-medulla projections whose activation enhances neuropathic hypersensitivity and whose inhibition reduces it [@zhang2023somatostatin]. The brainstem therefore regulates gain in both directions rather than operating as a one-way brake.
Local spinal circuits provide another form of control. Melzack and Wall’s gate-control theory proposed that dorsal-horn transmission is shaped by interactions among nociceptive afferents, low-threshold mechanoreceptive afferents, local interneurons, and descending signals [@melzack1965gate]. The original circuit has been revised, but its central insight remains: the dorsal horn of the spinal cord is an active computational site, not a passive relay. The local processing illustrated in Figure 22.2 is therefore part of pain regulation. Rubbing the skin around a minor injury can reduce pain partly because tactile input recruits spinal circuitry that competes with or inhibits nociceptive transmission.
Attention, expectation, threat, perceived control, and the availability of competing actions can all alter pain. Placebo analgesia provides one experimentally controlled demonstration of contextual modulation [@zunhammer2021placebo]. These effects need not imply an inner observer deciding whether an injury deserves attention. Cortical, hypothalamic, amygdalar, and brainstem circuits continually adjust sensory gain according to the organism’s state and circumstances.
Behavior closes an additional loop. Withdrawal removes tissue from the stimulus; guarding changes posture and movement; escape changes the environment; rest and treatment seeking change the body; and persistent avoidance can eventually produce weakness and deconditioning. The pain system does not merely estimate threat. It changes the conditions that will generate the next sensory sample.
23.6 When pain becomes uncoupled from immediate tissue threat
The adaptive logic of acute pain is clearest when a noxious event triggers withdrawal, guarding, and healing. Several clinical phenomena show that the relationship between injury and experience can become less direct. They do not make pain unreal. They reveal the many levels at which the system can be altered.
23.6.1 Referred pain
Pain can be felt at a location different from the tissue that generated the initiating signal. Myocardial ischemia, for example, often produces chest discomfort and may also be referred or radiate to the arm, shoulder, neck, or jaw.
The standard explanation emphasizes convergence. Visceral and somatic afferents can influence some of the same spinal projection neurons, creating ambiguity about the source of activity. Because somatic events are more common and spatially better mapped, the resulting sensation may be assigned to a region of the body surface associated with those neurons [@murray2009referred].
Convergence is a plausible substrate, but it is not a complete explanation. It does not by itself account for the consistency of some referral patterns, their variability across people, or the contributions of supraspinal processing. Referred pain is best treated as a localization problem produced by convergent and distributed circuitry rather than as evidence that the brain simply makes a categorical mistake at one synapse.
23.6.2 Sensitization: hyperalgesia and allodynia
Pain systems can become more responsive after tissue injury, inflammation, or damage to the nervous system. Hyperalgesia is an increased pain response to a stimulus that is normally painful. Allodynia is pain evoked by a stimulus that does not normally provoke pain, such as light brushing of the skin. These are clinical descriptions; neither term specifies a single mechanism.
Peripheral sensitization can lower the thresholds of nociceptors near an injury. Central sensitization refers to increased responsiveness within central nociceptive pathways. Repeated or intense input can strengthen excitatory transmission, enlarge receptive fields, increase intrinsic excitability, and reduce inhibition in dorsal-horn circuits. Low-threshold tactile input can then gain access to spinal output pathways that normally respond most strongly to noxious events.
Direct imaging of dorsal-horn populations in mice has shown this transformation at the circuit level. After capsaicin-induced sensitization, some spinal neurons developed responses to previously innocuous input, their receptive fields expanded, and spinoparabrachial output neurons became more responsive to low-threshold stimulation [@warwick2022central]. The experiment does not provide one universal mechanism for all allodynia, but it shows how touch can acquire access to a nociceptive output pathway without the gentle touch itself damaging tissue.
Sensitization is protective in the short term. A lowered threshold around an injury discourages contact and movement. The same plasticity becomes costly when it persists after its original protective purpose has passed.
23.6.3 When acute pain becomes chronic
Chronic pain is commonly defined as pain that persists or recurs for more than three months [@treede2019chronic]. The time criterion is useful, but chronic pain is not simply acute pain that has continued for longer. The biological system itself may change.
Some chronic pain is sustained by ongoing tissue disease or inflammation. Some is neuropathic, caused by a lesion or disease of the somatosensory nervous system. In other conditions, altered nociceptive processing is prominent even when ongoing tissue damage or a discrete neural lesion does not fully explain the experience. These mechanisms can coexist in the same person and can shift over time.
The term nociplastic pain has been proposed for pain in which altered nociception is prominent but ongoing tissue damage or a lesion or disease of the somatosensory system does not adequately account for the experience [@kosek2016nociplastic]. It is a mechanistic descriptor, not a single diagnosis or a claim that one mechanism has been identified. It should not be treated as synonymous with central sensitization, and nociceptive, neuropathic, and nociplastic mechanisms can coexist.
Persistent peripheral activity, spinal sensitization, altered inhibition, immune signaling, descending facilitation, learning, expectation, sleep disruption, stress, and changes in movement can all contribute. None of these factors means that chronic pain is “merely psychological.” Psychological and social variables alter neural and bodily systems, just as inflammation and nerve injury do. The relevant question is not whether pain is physical or psychological, but which interacting mechanisms are maintaining it in a particular case.
Viewed through the control architecture developed in this chapter, some chronic pain states can be understood as failures of a normally temporary protective state to disengage. Sensory gain remains elevated; ordinary bodily signals continue to receive high priority; guarding and avoidance persist; autonomic and defensive systems remain recruited; descending facilitation sustains spinal responsiveness; and reduced movement can generate weakness, stiffness, and new adverse bodily signals. This is not a claim that chronic pain is caused by a false belief. It is a claim that a distributed protective system can become self-maintaining at several levels.
This distinction matters for treatment. Rest and guarding may be adaptive immediately after injury but become harmful when they produce long-term deconditioning and avoidance. A drug that reduces peripheral inflammation may help one pain state and do little for another maintained principally by nerve injury or central sensitization. Chronic pain therefore requires explanation at several levels rather than the search for one hidden lesion or one cortical pain center.
23.6.4 Phantom pain without a limb
After amputation, many people continue to experience a phantom limb. The phantom may have a position and shape, may appear to move, and may produce non-painful sensations. In some people it is painful. Phantom-limb pain demonstrates that the experienced body is not identical to the current physical body, but it does not establish one mechanism.
Peripheral activity can remain important. Injured nerves and neuromas in the residual limb can generate abnormal discharges. A multicenter randomized trial found that a six-day continuous peripheral nerve block reduced established phantom pain four weeks after treatment relative to placebo [@ilfeld2021phantom]. The missing limb therefore does not imply the absence of nociceptive input from the remaining peripheral nervous system.
The preceding chapter’s discussion of cortical-map plasticity also matters here. Established amputees can retain organized activity associated with the missing hand, and longitudinal observations after planned amputation found stable hand and lip representations rather than large-scale invasion of the hand territory by the face [@makin2017stability; @makin2023against; @makin2025stable]. The older claim that phantom pain is generally caused by neighboring body parts taking over the missing limb’s cortical territory is therefore not secure. Peripheral ectopic activity, residual-limb pathology, spinal and thalamic changes, and persistent cortical body representations may each contribute, with different weight across individuals.
23.6.5 Congenital insensitivity to pain
The reverse dissociation is equally revealing. Several rare genetic conditions produce profound insensitivity to pain. One well-studied form is caused by loss-of-function mutations in SCN9A, the gene encoding the voltage-gated sodium channel NaV1.7 [@cox2006scn9a]. Without normal NaV1.7 function, nociceptive neurons have great difficulty initiating or propagating signals that ordinarily support pain.
The condition is not a benefit. Affected people may accumulate burns, wounds, fractures, tongue and lip injuries, infections, and joint damage because injuries do not trigger normal withdrawal, guarding, or help-seeking. Pain is aversive because aversion is part of its adaptive function. A body unable to assign sufficient urgency to injury is poorly protected from repeated harm.
Many people with SCN9A-related pain insensitivity are also anosmic. NaV1.7 is required not only in nociceptive pathways but also for effective transmission from olfactory sensory neurons [@weiss2011scn9a]. The pairing of pain insensitivity and anosmia therefore provided an important clue that one ion channel participates in two apparently unrelated sensory systems.
The comparison between chronic pain and congenital pain insensitivity exposes the control problem. Too much gain can make protection persist after it is useful; too little can leave the body undefended. The biological task is not to eliminate pain but to regulate it in proportion to threat, competing demands, and recovery.
23.7 Looking ahead
Temperature and pain begin with peripheral receptors and afferent fibers, but their central organization is distributed. Thermal and nociceptive signals are transformed in the spinal cord and enter overlapping systems that localize a threat, estimate bodily condition, assign priority, organize defense and learning, and regulate the gain of the next signal. Fast and slow afferents provide different temporal components, but no single fiber class, thalamic nucleus, cortical area, or brainstem circuit contains pain.
Sensitization, chronic pain, and phantom pain show how an adaptive control system can become persistent or partly uncoupled from its original trigger. Congenital pain insensitivity shows the opposite failure: without sufficient aversive signaling, the body cannot protect itself. Pain is therefore neither a simple damage meter nor an arbitrary construction. It is a regulated biological state in which sensation, valuation, bodily condition, learning, and action are inseparable.
The unit now turns toward hearing and vision. These distance senses extend the spatial and temporal horizon of control, allowing events to guide behavior before they reach the body. The same principle will remain in force: sensory systems do not merely describe the world; they extract the information needed to act within it.
Reasonably settled:
- Pain and nociception are distinct. Pain cannot be inferred solely from activity in sensory neurons.
- Innocuous warming and cooling are genuine sensations, not merely weaker versions of thermal pain; increasingly extreme temperatures recruit high-threshold afferents and nociceptors.
- Nociceptors directly transduce high-threshold mechanical, thermal, and chemical events, while inflammatory mediators can activate or sensitize their endings.
- A brief cutaneous noxious stimulus can produce Aδ-mediated first pain followed by C-fiber-mediated second pain.
- Nociceptive traffic fans into overlapping discriminative-sensorimotor, body-state, priority-setting, autonomic, learning, and defensive systems rather than terminating in a single pain center.
- Posterior insular-opercular cortex participates importantly in thermal and nociceptive processing, while cingulate systems contribute to urgency, anticipation, aversion, and action allocation.
- Parabrachial, amygdalar, hypothalamic, and PAG circuits link nociceptive input to defensive behavior, autonomic regulation, learning, and descending control; the finest cell-type evidence comes mainly from rodents.
- Nociceptive processing is regulated at peripheral, spinal, brainstem, thalamic, and cortical levels. Descending systems can inhibit or facilitate transmission.
- Peripheral and central sensitization can produce hyperalgesia and allodynia.
- Loss-of-function mutations in SCN9A can produce profound pain insensitivity and anosmia, revealing the adaptive necessity of pain.
Genuinely unsettled, and presented as such:
- Why some acute pain resolves while other pain becomes chronic. Multiple peripheral, spinal, immune, descending, cognitive, and behavioral factors contribute, but their causal weight differs among conditions and individuals.
- The mechanisms grouped under nociplastic pain. The term identifies a pattern of altered nociception and an explanatory gap; it does not specify one common mechanism.
- The complete mechanism of referred pain. Somatic-visceral convergence is important but does not fully explain stable referral patterns and individual variability.
- The mechanisms of phantom-limb pain. Large-scale cortical takeover is not a secure general explanation; peripheral and central contributions probably vary across people.
- The exact thalamic routes into human posterior insular and cingulate cortex. VMpo remains disputed, spinothalamic terminations are widespread, and direct thalamocortical and corticocortical routes probably make different contributions at different times.
- The precise division of labor among posterior insula, operculum, S1, S2, anterior insula, and anterior/midcingulate cortex. These regions make differentiated contributions but interact within recurrent networks rather than forming a simple serial chain.
- How directly rodent parabrachial–amygdala–hypothalamus–PAG cell types map onto humans. The broad architecture is supported, but the fine correspondence remains uncertain.
The durable lesson is not “two pains, two pathways, two destinations.” It is that thermal and nociceptive signals enter a distributed control architecture that locates threat, estimates bodily condition, assigns priority, organizes defense and learning, regulates gain, and changes the action that determines what signal arrives next. The system is adaptive precisely because it is modifiable—and vulnerable precisely for the same reason.