28 Taste: Chemicals at the Threshold of the Body
Receptors, Flavor, and Ingestive Control
Taste is chemosensation at the threshold of ingestion. Its receptors sample chemicals that have already entered the mouth but have not yet been swallowed, placing the system close to the boundary between exteroception and interoception. On the organizing axes introduced in Chapter 21, taste is an extremely proximal and largely reactive sense: it buys little lead time, but sometimes enough to alter ingestion, initiate protective responses, and prepare the digestive system for what may follow.
The chemicals on the tongue are still outside the body in the physiological sense. Taste is therefore not simply interoception in disguise. What makes it unusual is the speed with which information about an external substance can be brought into contact with ongoing bodily needs. Salt can become more attractive during sodium depletion; the appeal of a familiar food can decline during a meal; and a taste that once preceded illness can later provoke rejection. Taste quality and taste value are consequently coupled, but they are not the same thing.
The chapter follows the same route used in the somatosensation chapter (Chapter 22), moving from stimulus and receptor organ to first-order sensory neuron, first central processor, thalamus, cortex, and action. Taste modifies the canonical plan in an instructive way. The receptor cell is not itself the first-order neuron, the first central processing stage lies in a brainstem complex closely involved in ingestion and visceral control, and the cortical representation does not appear to contain the clean map once proposed for it.
28.1 Taste is not flavor
In ordinary speech, taste often means the entire experience of food. In sensory physiology, the term is narrower. Taste is the signal produced when water-soluble chemicals act on receptor cells in taste buds. The perceived flavor of a food combines that signal with several others:
- volatile molecules released in the mouth reach the olfactory epithelium by the retronasal route;
- somatosensory receptors report texture, viscosity, temperature, pressure, and movement;
- nociceptive and thermosensitive afferents contribute chemesthesis—the burn of capsaicin, the cooling of menthol, the sting of carbonation, and some forms of astringency;
- expectations and learned associations help identify the substance and predict its consequences.
This is why food seems to lose much of its “taste” during nasal congestion even when sweet, salty, sour, bitter, and umami can still be detected. Much of what has disappeared is retronasal smell. Conversely, a chili pepper can remain painfully hot even when its gustatory qualities are weak, because capsaicin activates trigeminal nociceptors rather than a taste receptor. The mouth is therefore not served by a single food sense. Taste, smell, touch, temperature, and chemesthesis enter by different peripheral routes and are progressively combined into an experience attributed to the food in the mouth.
Recent human imaging makes the distinction especially clear. Tastants strongly engage bilateral dorsal mid-insula, while retronasal odors engage olfactory cortex. After tastes and odors have been learned as parts of the same flavor, however, their activity patterns overlap in more anterior and ventral insular territory. Flavor is thus integrated from initially distinct signals rather than being a synonym for taste [@khorisantono2025flavour].
The distinction will matter again in the clinical section. A patient who says that food no longer “tastes right” may have lost basic taste, smell, flavor integration, oral somatosensation, or some combination of them. The pathway must be examined before the complaint can be localized.
28.2 Taste buds sample dissolved chemicals
28.2.1 Taste buds are sensory organs
Taste receptors are housed in taste buds, compact sensory organs found in fungiform papillae toward the front of the tongue, foliate papillae along its sides, circumvallate papillae near its rear, and smaller populations in the palate and oropharynx. The numerous filiform papillae that give the tongue much of its rough surface do not contain taste buds; they serve principally mechanical and somatosensory functions. The familiar schoolroom “tongue map,” with sharply separated sweet, salty, sour, and bitter zones, is therefore wrong. All established qualities can be detected across taste-bearing regions, although regional sensitivity is not perfectly uniform.
A taste bud contains elongated epithelial cells whose apical microvilli reach the oral cavity through a taste pore. Dissolved chemicals contact those apical membranes. The cells are not permanent neurons. They arise from the surrounding epithelium and are continuously replaced, so gustatory afferents must maintain or renew functional contacts as receptor cells turn over [@beidler1965renewal].
Several cell classes participate. Type II receptor cells contain most of the molecular machinery for sweet, bitter, and umami. Type III cells contain the principal sour-transduction machinery and form conventional chemical synapses with afferent endings. Type I cells perform support functions and may also contribute to some forms of salt processing. These categories are useful, but they should not be mistaken for five perfectly isolated cellular pipes. Salt uses more than one pathway, and interactions among neighboring cells help transmit the final signal out of the taste bud.
This arrangement introduces an important variation on the canonical sensory route. In touch, the receptor ending is part of the first-order sensory neuron. In taste, an epithelial receptor cell transduces the chemical stimulus and then communicates with the peripheral ending of a separate cranial sensory neuron. The sequence is:
sapid chemical → taste-bud cell → transmitter release → first-order cranial afferent
28.2.2 Five established qualities, several transduction strategies
The five conventionally established taste qualities do not use one common receptor mechanism. Sweet, umami, and bitter begin with G-protein-coupled receptors; sour and at least one form of salty taste begin with ion channels. The central distinctions are summarized in Table 28.1.
| Quality | Principal peripheral mechanism | Important qualification |
|---|---|---|
| Sweet | T1R2–T1R3 receptor heterodimer | Sweetness is associated with many carbohydrates but is not a direct measurement of caloric value. |
| Umami | T1R1–T1R3 receptor heterodimer, enhanced by nucleotides | Principally signals free glutamate and related compounds rather than “protein” as a whole. |
| Bitter | A family of roughly 25 functional human T2R receptors | Many toxins are bitter, but bitterness is neither necessary nor sufficient for toxicity. |
| Sour | Proton detection centered on OTOP1 in Type III cells | The strongest causal molecular evidence comes from mouse taste cells, nerves, and behavior. |
| Salty | ENaC-dependent low-sodium pathway in rodents, plus additional pathways | Human salt transduction is heterogeneous and is not adequately described by ENaC alone. |
The sweet receptor is a partnership between T1R2 and T1R3; the related T1R1–T1R3 heterodimer contributes to umami [@nelson2001sweet; @nelson2002umami; @li2002umami]. Bitter compounds are detected by the larger T2R family [@adler2000t2r; @chandrashekar2000t2r]. Some T2Rs respond to a relatively restricted set of compounds, whereas others are more broadly responsive [@meyerhof2010bitter]. The contrast between one principal sweet-receptor heterodimer and a larger bitter-receptor family is consistent with the much greater chemical diversity of bitter ligands. It should not, however, be turned into a simple equation in which sweetness means nutrition and bitterness means poison. Evolution has supplied initial biases, not an infallible food assay.
Sweet, bitter, and umami receptors feed into substantially shared intracellular machinery. Receptor activation engages a G protein, often including gustducin, and the PLCβ2–IP3R3 pathway releases intracellular calcium. The calcium-activated cation channels TRPM4 and TRPM5 then help depolarize the cell. Removing either TRPM channel impairs these tastes in mice; removing both produces a much more complete loss than deletion of TRPM5 alone [@duttabanik2018trpm]. The older statement that a TRPM5 knockout simply abolishes all sweet, bitter, and umami perception is therefore too absolute.
The output of a Type II cell is unusual. These cells lack the conventional synaptic vesicles found at most chemical synapses. Instead, depolarization opens an ATP-permeable channel formed by CALHM1 and CALHM3, releasing ATP onto purinergic receptors on the gustatory afferent ending [@taruno2013calhm1; @ma2018calhm3]. This is sometimes called a channel synapse: the same membrane channel provides the route by which the transmitter leaves the receptor cell.
ATP signaling is a common link between taste buds and gustatory nerves. The afferent endings express P2X2 and P2X3 receptors, and a topical P2X2/P2X3 antagonist recently suppressed sweet, salty, sour, bitter, and savory tastes in human participants while sparing oral tingle and astringency [@flammer2024p2x]. The result draws a sharp functional boundary between gustation and nongustatory oral sensation. It does not imply that every taste-bud cell releases ATP in the same way: Type III cells make conventional synapses and use additional transmitters, so sour reaches the common purinergic output through a more complex local circuit.
Sour transduction illustrates how recently some basic mechanisms were identified. Acids supply protons, and the proton channel OTOP1 is expressed in Type III cells. Eliminating OTOP1 in mice strongly reduces the proton current in those cells, gustatory-nerve responses to acids, and normal sour-guided behavior [@tu2018otop1; @teng2019otop1]. OTOP1 therefore provides the principal molecular entry point for sour, although acidification also changes other membrane conductances within the receptor cell.
Salt is less tidy. In mice, an amiloride-sensitive pathway built around the epithelial sodium channel ENaC detects relatively low sodium concentrations and contributes to sodium attraction [@chandrashekar2010salt; @ohmoto2020sodium]. Higher salt concentrations additionally recruit amiloride-insensitive pathways, including subsets of Type III and bitter-responsive cells, and become aversive [@oka2013highsalt]. Human psychophysical effects of amiloride are variable and often incomplete. Some experiments found no reduction in NaCl saltiness, even with localized stimulation, whereas others found modest reductions in total intensity or in sour side tastes more than in saltiness itself [@desor1989amiloride; @ossebaard1996amiloride]. ENaC is consequently a well-established component of rodent sodium taste, not a complete settled account of human salty taste.
A candidate taste requires more than a molecule that can be detected in the mouth. A strong case would include a distinguishable oral percept not reducible to the established tastes, smell, texture, or irritation; a transduction mechanism; a route into gustatory afferents; and reproducible behavioral or perceptual consequences.
Free fatty acids meet several of these criteria and can produce a distinguishable human percept sometimes called oleogustus, although fat-rich foods also carry powerful texture and odor signals [@running2015oleogustus]. Humans can detect some glucose oligomers even when the canonical sweet receptor is blocked, suggesting a carbohydrate-sensitive mechanism beyond ordinary sweetness [@pullicin2017glucose]. OTOP1 also mediates much of the neural and behavioral response to ammonium chloride in mice [@liang2023ammonium]. Each finding expands the biology of oral chemosensation. None by itself requires replacing “five established tastes” with a universally accepted new number.
28.3 The brainstem is taste’s first sensory processor
28.3.1 Three cranial routes, three sensory ganglia
Taste-bud cells communicate with peripheral endings carried by three cranial nerves. Branches of the facial nerve (VII) supply taste buds on the anterior tongue and palate; the glossopharyngeal nerve (IX) supplies much of the posterior tongue; and the vagus nerve (X) supplies taste buds in the epiglottic and upper pharyngeal region. These territories overlap at their borders and should not be treated as perfectly partitioned thirds.
The first-order neuron is again distinct from the receptor cell. Its cell body lies in a cranial sensory ganglion:
- facial gustatory afferents have their somata in the geniculate ganglion;
- glossopharyngeal afferents have their somata in the petrosal ganglion;
- vagal gustatory afferents have their somata in the nodose ganglion.
Each neuron extends a peripheral process to taste buds and a central process into the medulla. This is the cranial counterpart of the dorsal-root-ganglion organization introduced in Chapter 22, although the peripheral receptor organ is different.
The central processes terminate principally in the rostral nucleus of the solitary tract, or rostral NTS. The distinction between rostral and caudal matters. Much vagal information from the thoracic and abdominal organs terminates farther caudally in the NTS, whereas oral gustatory input is concentrated toward its rostral end. Taste therefore enters the same elongated nuclear complex that participates in visceral regulation, but not an anatomically undifferentiated pool in which every taste signal is immediately fused with blood pressure, gastric distension, and other visceral variables.
28.3.2 An early sensory and ingestive hub
Like the spinal cord in Chapter 22, the NTS is not merely a cable junction on the way to awareness. It begins processing the signal. Recordings from awake, freely licking rats reveal taste-responsive neurons that are usually responsive to more than one quality, neurons synchronized to licking, and activity that predicts the beginning or end of a lick bout. Spike timing can carry information beyond total spike count [@roussin2012nts]. Taste in the NTS is therefore already embedded in the sensorimotor act by which the animal samples and ingests a liquid.
The rostral NTS also distributes information into parallel routes. Ascending neurons contribute to forebrain perception and evaluation, while other populations project within the brainstem to circuits involved in salivation, licking, swallowing, and rejection. Anatomical tracing indicates that major ascending and descending projections arise from substantially separate NTS populations rather than from one homogeneous relay [@halsell1996ascending]. Descending signals from forebrain structures can in turn alter taste-responsive activity in the brainstem. The pathway is recurrent and parallel, in keeping with the general organization developed in Chapter 21.
This is the appropriate anatomical connection to the interoceptive material from Unit II. Taste enters a brainstem complex positioned to coordinate oral chemical information with consummatory behavior and bodily state. The arrangement permits early interaction; it does not erase taste quality or turn gustation into a generic visceral signal. A useful formulation is that taste is contact exteroception delivered directly into an ingestive control system.
Many of the most elegant modern taste experiments are performed in mice, but the ascending pathway differs between rodents and primates.
In rodents, the principal route from rostral NTS passes through an obligatory gustatory relay in the parabrachial nucleus before reaching thalamus. The parabrachial region also sends prominent projections to hypothalamic and amygdalar circuits. In macaques, by contrast, NTS neurons send a substantial direct projection to the parvocellular gustatory region of thalamus, without the same obligatory pontine relay [@norgren1971taste; @beckstead1980monkey].
Mouse circuit results remain highly informative, but a diagram traced in mouse cannot simply be relabeled as the human pathway.

28.4 Vomiting is a centrally patterned defensive act
Taste can change the fate of a substance while it remains in the mouth: sampling can stop, the substance can be expelled, and swallowing can be withheld. Once material has passed that oral threshold, defense requires a different scale of coordination. Vomiting removes material that has entered the upper digestive tract, but it is not simply an extreme response to bitterness. Gastrointestinal signals, blood-borne chemicals, vestibular input, and learned or anticipatory cues can all recruit the same organized act. Three terms should be kept separate: nausea is the consciously experienced state of visceral malaise, retching is the repeated forceful motor pattern that prepares for expulsion, and vomiting (emesis) is the expulsive event itself. Nausea need not culminate in vomiting, and the motor pattern can be studied even when nothing is expelled.
28.4.1 Detecting a threat is not generating the act
Chemicals or irritation in the stomach and intestine recruit vagal afferents terminating in the caudal NTS (nucleus of the solitary tract) and adjoining dorsal vagal complex. The area postrema, one of the sensory circumventricular organs introduced in Unit II, samples blood and cerebrospinal fluid without an ordinary blood–brain barrier and is therefore positioned to detect circulating emetic agents. Vestibular nuclei provide another route, and forebrain signals can recruit the system when a sight, smell, or context has previously predicted illness.
The area postrema is consequently important, but it is not the vomiting center. Lesioning it prevents emesis produced by many blood-borne drugs while sparing vomiting triggered by vestibular stimulation and by direct activation of abdominal vagal afferents [@miller1994area]. Its role is detection and recruitment within a larger system. Note also that gustatory afferents terminate principally in rostral NTS, whereas visceral signals capable of initiating emesis engage more caudal NTS: taste and vomiting are positioned to interact without constituting one undifferentiated pathway.
28.4.2 Fictive vomiting reveals a central pattern generator
The strongest evidence for a central pattern generator comes from decerebrate, paralyzed dogs. Stimulating vagal afferents produced coordinated bursts in the phrenic and abdominal motor nerves even though the animal could neither execute the movements nor return movement-generated sensory feedback. This centrally generated output is called fictive vomiting. Destroying a reticular region dorsomedial to the retrofacial nucleus abolished the patterned response, and neurons there project directly to respiratory premotor neurons in the caudal ventral respiratory group [@koga1998vomiting; @koga1997descending].
A central pattern generator need not be a clock that oscillates continuously. It is a network that, once recruited, imposes an ordered temporal pattern on multiple motor populations without depending on each movement to cue the next. The older phrase vomiting center therefore encourages too simple a picture: the experiments identify an essential medullary reticular region, but the complete act depends on interactions among the dorsal vagal complex, reticular pattern-generating neurons, respiratory premotor groups, pharyngeal and laryngeal motor systems, vagal output to the gut, and spinal motor neurons. A distributed emetic pattern-generating network is the more accurate formulation.
28.4.3 A motor ballet
No single muscle or organ produces vomiting. Before retching begins, the proximal stomach and lower esophageal sphincter relax and the pharynx and upper esophagus adopt a protective configuration. During retching, precisely phased activity in the diaphragm and abdominal muscles generates large pressure changes while the airway is protected. Expulsion requires yet another configuration: the gastroesophageal route opens, abdominal pressure rises, and laryngeal closure keeps material out of the airway [@lang1986gastrointestinal; @lang1993vomiting; @lang2002airway]. Even the diaphragm divides its labor — the costal portion largely retains its respiratory activation pattern while the crural segment at the esophageal hiatus is reconfigured in concert with opening the gastroesophageal passage [@park2025emesis]. Successful expulsion depends on selective recruitment and inhibition, not on a global contraction of everything at once.
Vomiting thus reveals a general control architecture: several sensory routes can trigger it, a distributed brainstem network organizes its sequence, and autonomic and somatic effectors are temporarily reorganized into a single defensive act.
28.5 From thalamus to gustatory cortex
In primates, ascending NTS output reaches a parvocellular gustatory region of the ventral posterior medial thalamus, commonly called VPMpc. Its position near the VPM territory for facial somatosensation is anatomically convenient but should not imply that the two systems are interchangeable. VPMpc receives gustatory input and projects prominently to insular and opercular cortex [@pritchard1986monkey].
The thalamic relay should be read through the account in Section 21.3. It is neither a passive way station nor a place at which taste waits unchanged for cortex. Thalamic neurons participate in recurrent loops with cortex, and the activity passing through them is shaped by behavioral state, attention, and descending input. At the same time, taste-related signals branch toward structures involved in motivation, autonomic control, and learned value. The familiar receptor → brainstem → thalamus → cortex sequence is therefore the main ascending road, not the entire network.
Human imaging consistently identifies taste-responsive cortex in the dorsal mid-insula and adjoining frontal or parietal opercular territory [@avery2020taste]. More anterior and ventral insular regions participate in combining taste with retronasal odor and other information about flavor [@khorisantono2025flavour]. Orbitofrontal cortex, striatum, amygdala, hypothalamic systems, and somatosensory cortex contribute additional information about reward, expectation, bodily state, texture, and action. Referring to all of this as “the insula” can conceal meaningful anatomical differences. Primary gustatory responses, flavor integration, interoception, and valuation engage overlapping but nonidentical insular territories and broader networks.
This distributed arrangement also prevents a misleading analogy with the pain chapter (Chapter 23). Taste and pain both recruit insular cortex, as do many interoceptive and oral signals, but shared cortical territory does not make them one function. The useful principle is convergence: signals relevant to the body’s condition and possible action interact in insular networks. The claim that one insular region simply attaches “good” or “bad” to every sensation is much stronger than the evidence permits.
28.6 How taste quality is encoded
28.6.1 Selective peripheral channels, mixed central populations
The traditional coding debate contrasts labeled lines, in which narrowly tuned neurons carry distinct qualities, with across-fiber patterns, in which quality is represented by activity across broadly tuned populations. Taste biology suggests that the alternatives apply differently at different stages rather than competing as complete descriptions of the entire pathway.
At the receptor-cell level, there is substantial selectivity. Sweet-, bitter-, and umami-responsive Type II cells express different receptor families, and manipulating those cell populations can strongly and selectively alter taste-guided behavior [@zhang2003coding]. Sour depends heavily on OTOP1-expressing Type III cells, while low-sodium taste in mice uses a distinguishable ENaC-dependent population. The periphery therefore begins with more channel specificity than older versions of a wholly distributed account allowed.
Selectivity becomes less exclusive centrally. In awake rats, most taste-responsive NTS neurons respond to several qualities [@roussin2012nts]. In awake mice, about two thirds of recorded gustatory-cortical neurons responded differentially to three or four tested tastes. Ensemble activity first carried information about taste identity and later—beginning roughly half a second after delivery—shifted toward palatability-related activity [@levitan2019taste]. Which neurons fire, how strongly they fire, and how the population changes through time can all contribute information.
The most useful synthesis is therefore hierarchical:
relatively selective peripheral channels converge into mixed, dynamic central populations.
A population code and a temporal code are not mutually exclusive. Temporal coding is a population pattern considered across time. Nor does a broadly responsive central neuron imply that the peripheral distinctions were imaginary. Convergence allows the nervous system to compare qualities, concentrations, oral movements, learned context, and current bodily state while retaining enough information to discriminate the stimulus.
28.6.2 Does gustatory cortex contain a taste map?
The map question follows naturally from the unit overview. Vision preserves position on the retina, somatosensation preserves position on the body, and audition preserves an ordered frequency axis. An influential 2011 mouse study reported spatially segregated cortical hotspots for sweet, bitter, salty, and umami, suggesting an analogous gustotopic map [@chen2011gustotopic].
Subsequent work has not supported that strong canonical picture. Imaging and electrophysiology in awake mice have generally found taste-responsive neurons to be sparse, overlapping, broadly tuned, and distributed across gustatory cortex rather than confined to reproducible quality-specific territories [@chen2021distributed; @fletcher2017overlapping; @levitan2019taste]. Human 7-Tesla imaging likewise found that sweet, salty, and sour all activated bilateral dorsal mid-insula. Taste identity could be decoded from the local multivoxel pattern, but no cortical location consistently preferred one quality across participants [@avery2020taste].
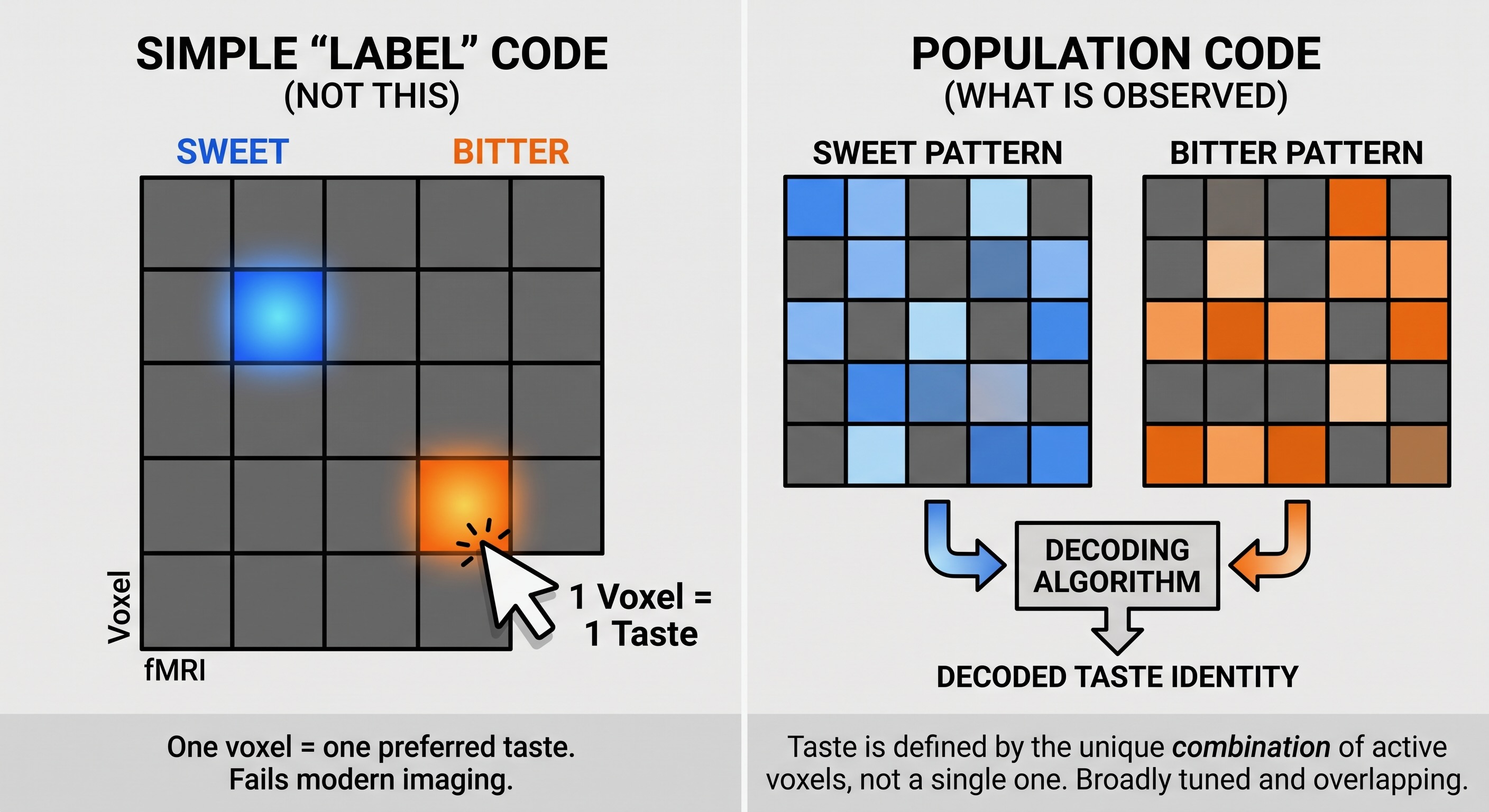
A small 2026 high-field study adds a useful qualification. All five qualities could be decoded from local insular clusters within individuals, but cluster locations varied markedly from person to person. Valence was decoded less strongly and from clusters with relatively little overlap with the quality patterns. With only eight participants, the result is preliminary, but it fits an individually variable population code better than a fixed anatomical atlas [@canna2026taste].
The present conclusion is therefore more precise than “taste has no spatial organization.” Local biases and clusters may exist, and spatial patterns contain information. What has not emerged is a conserved cortical map with a stable sweet area, bitter area, sour area, and so forth. One likely reason is that the established taste qualities are category-like and are not laid out along an ordered peripheral sheet comparable to retinal location, body surface, or cochlear frequency. That explanation is plausible rather than proven, but it connects the negative gustotopy evidence to the broader question of what a sensory map is a map of.
28.7 Taste quality is not taste value
Taste signals arrive with powerful behavioral biases. Newborn mammals need not learn from first principles that concentrated quinine is ordinarily rejected or that sweetness can support ingestion. But innate bias is not a fixed verdict. Value changes with concentration, physiological condition, learning, expectation, and post-ingestive outcome.
Salt makes the point especially clearly. Low concentrations can be attractive, high concentrations are ordinarily aversive, and sodium depletion can make animals tolerate or seek salt that they would otherwise reject. Mouse experiments have identified partly separate circuits for sodium appetite and for state-dependent tolerance of aversive salt, showing that internal state changes both the drive toward sodium and the sensory barrier that limits its intake [@zhang2023sodiumvalence]. A taste quality can therefore remain identifiable while its behavioral value changes.
The cortical time course points in the same direction. In awake mice, identity-related ensemble activity emerges before palatability-related activity [@levitan2019taste]. Human high-field data likewise suggest that quality and rated valence can be decoded from partly distinct and individually variable insular patterns, although the current sample is too small for a strong anatomical conclusion [@canna2026taste]. Quality and value interact, but neither is reducible to the other.
A particularly informative mouse experiment traced sweet- and bitter-responsive gustatory-cortical populations to different targets in the basolateral amygdala. Manipulating those projections altered appetitive and aversive behavior, and silencing the amygdala reduced the normal behavioral pull of sweet and bitter while sparing discrimination between them [@wang2018valence]. The experiment provides strong causal evidence that taste identity and behavioral value can be partly dissociated in a mouse circuit.
It does not show that cortex represents only identity, that the amygdala alone assigns value, or that the segregated projections constitute a general “map of good and bad.” Palatability-related activity is already present in gustatory cortex, and brainstem circuits influence acceptance and rejection before cortical processing is complete. Taste value is best understood as a distributed, state-dependent control variable assembled through interactions among sensory, visceral, learning, and action systems.
28.8 When taste is lost or distorted
Clinical complaints distinguish several forms of dysfunction. Ageusia is the loss of taste, hypogeusia a reduction in taste sensitivity, and dysgeusia a distorted or persistently unpleasant taste. These should be separated from loss of smell and from broader disturbances of flavor, even though patients may use the same everyday language for all of them.
Cortical stroke can produce severe gustatory disturbance. One well-described patient was a 75-year-old woman whose left middle cerebral artery stroke involved the insula. After returning to her usual diet, she reported that “everything tastes like dirt.” Eating became unpleasant, social meals lost their appeal, and she lost about 14 pounds [@dutta2013stroke]. The case demonstrates that taste and flavor disturbance can threaten nutrition and quality of life; it should not be dismissed as a minor sensory inconvenience.
The case does not provide a clean experiment in which chemical detection survived but hedonic meaning alone was removed. The patient initially had a larger neurological syndrome, experienced changes extending beyond food, and did not receive formal testing that cleanly separated basic taste from smell and flavor. More systematic lesion work shows that unilateral insular damage can impair taste discrimination, quality judgment, and naming on both sides of the tongue, with additional lateralized effects on some hedonic judgments [@stevenson2013lateralization]. The safest conclusion is that insular–opercular cortex is an important part of the network required for normal taste perception and interpretation, not that a single insular site constructs all gustatory experience or adds its value after the fact.
28.9 Looking ahead: from contact to distance
Taste occupies the contact end of the unit’s reactive–predictive gradient. A substance must already be in the mouth, but the system can still influence whether sampling continues, whether swallowing is initiated, and how digestive and motivational circuits respond. Its small amount of lead time is consequential because the next act may admit the substance more fully into the body.
Olfaction extends chemical sensing outward. Volatile molecules can be detected before an object is touched or ingested, allowing chemical information to guide approach, avoidance, search, and recognition at a greater distance. During eating, the retronasal route then joins odor with taste and oral somatosensation to create flavor. Olfactory information reaches primary olfactory cortex without the obligatory thalamic relay that precedes primary cortical processing in the other major sensory systems, although thalamic circuits participate in later attention, identification, and cortical interaction.
Taste has therefore supplied a compact variation on the shared sensory plan. Epithelial receptor cells transduce a restricted set of chemical properties; cranial sensory neurons carry the signal to a brainstem processor already engaged in ingestion; ascending pathways reach thalamic and insular–opercular networks; and quality, value, bodily state, and action become progressively intertwined without becoming the same variable. The next chapter asks how much earlier a chemical signal can begin to control behavior when it is carried through the air.
Reasonably settled:
- Taste is contact chemosensation at the threshold of ingestion. It is distinct from flavor, which also depends on retronasal olfaction, oral somatosensation, temperature, and chemesthesis.
- Taste-bud receptor cells are specialized epithelial cells rather than first-order neurons. Sweet, bitter, and umami use T1R or T2R GPCR pathways; OTOP1 is central to sour transduction; and ATP acting on P2X2/P2X3-bearing afferents is a crucial output signal from taste buds.
- Facial, glossopharyngeal, and vagal gustatory afferents have first-order somata in the geniculate, petrosal, and nodose ganglia and terminate principally in rostral NTS.
- Vomiting is not produced by a single medullary “center.” Several trigger routes recruit a distributed emetic pattern-generating network that coordinates autonomic and somatic effectors; the area postrema is one blood-facing trigger structure within that system.
- The main primate ascending route passes from NTS to the parvocellular gustatory thalamus and then to insular–opercular cortex. Rodents add an obligatory parabrachial relay.
- The traditional tongue-zone map is false. Likewise, current evidence does not support a fixed cortical atlas containing reproducible territories for each basic taste quality.
- Taste quality and taste value are coupled but separable. Both are represented across distributed, interacting circuits and can change with concentration, learning, and physiological state.
Still being refined:
- The full molecular basis of salty taste in humans. ENaC has a well-established role in rodent low-sodium taste, but human salt perception depends on additional and incompletely identified mechanisms.
- How many basic tastes should be recognized. Free fatty acids, glucose oligomers, ammonium, and other candidates satisfy some criteria, but no single evidential rule determines when a new oral chemosensory channel becomes a new canonical taste.
- How peripheral specificity becomes central population activity. Selective receptor-cell channels, convergence, response magnitude, and timing all contribute, but their relative importance changes across stages and behavioral conditions.
- The fine spatial organization of human gustatory cortex. Taste quality is decodable from insular patterns, yet those patterns appear overlapping and individually variable rather than chemotopically fixed.
- How taste is combined with bodily state and learned outcome. The relevant interactions begin in brainstem circuits and continue through thalamic, insular, amygdalar, orbitofrontal, striatal, and hypothalamic networks.
- The cellular organization of the human emetic pattern generator. The decisive central-pattern evidence comes from vomiting-capable species, whereas common laboratory rats and mice do not perform the complete act.