As described in Chapter 28, taste samples chemicals after they have entered the mouth. Olfaction samples chemicals before direct contact with their source. Airborne molecules can announce food, smoke, a familiar person, a predator, or a changing environment while there is still time to approach, avoid, investigate, or prepare. On the organizing axes introduced in Chapter 21, olfaction is therefore an exteroceptive distance sense. Its temporal reach is usually shorter and less geometrically precise than vision or audition, but it can nevertheless turn a chemical event elsewhere in the environment into information for action.
The pathway follows a recognizable sensory sequence. Molecules carried into the nasal cavity activate receptor-bearing olfactory sensory neurons in the olfactory epithelium. Those cells are both receptors and first-order neurons. Their axons enter the olfactory bulb, the system’s first central processor, where neurons expressing the same receptor identity converge onto glomeruli. Mitral and tufted cells then distribute bulbar output to several primary olfactory areas, including piriform cortex. Only after this first cortical stage does an important route pass through mediodorsal thalamus toward orbitofrontal cortex.
airborne chemical → olfactory sensory neuron → olfactory bulb → primary olfactory cortex → later thalamic and cortical processing
Olfaction thus modifies the canonical route from receptor to cortex without abandoning its logic. The receptor cell is itself the first-order neuron; the initial central pathway is predominantly ipsilateral; primary olfactory cortex is reached without an obligatory preceding thalamic relay; and its principal cortical target is three-layered paleocortex rather than six-layered sensory neocortex with a granular layer 4. These are specific architectural differences, not evidence that smell operates outside the general principles developed in the unit overview.
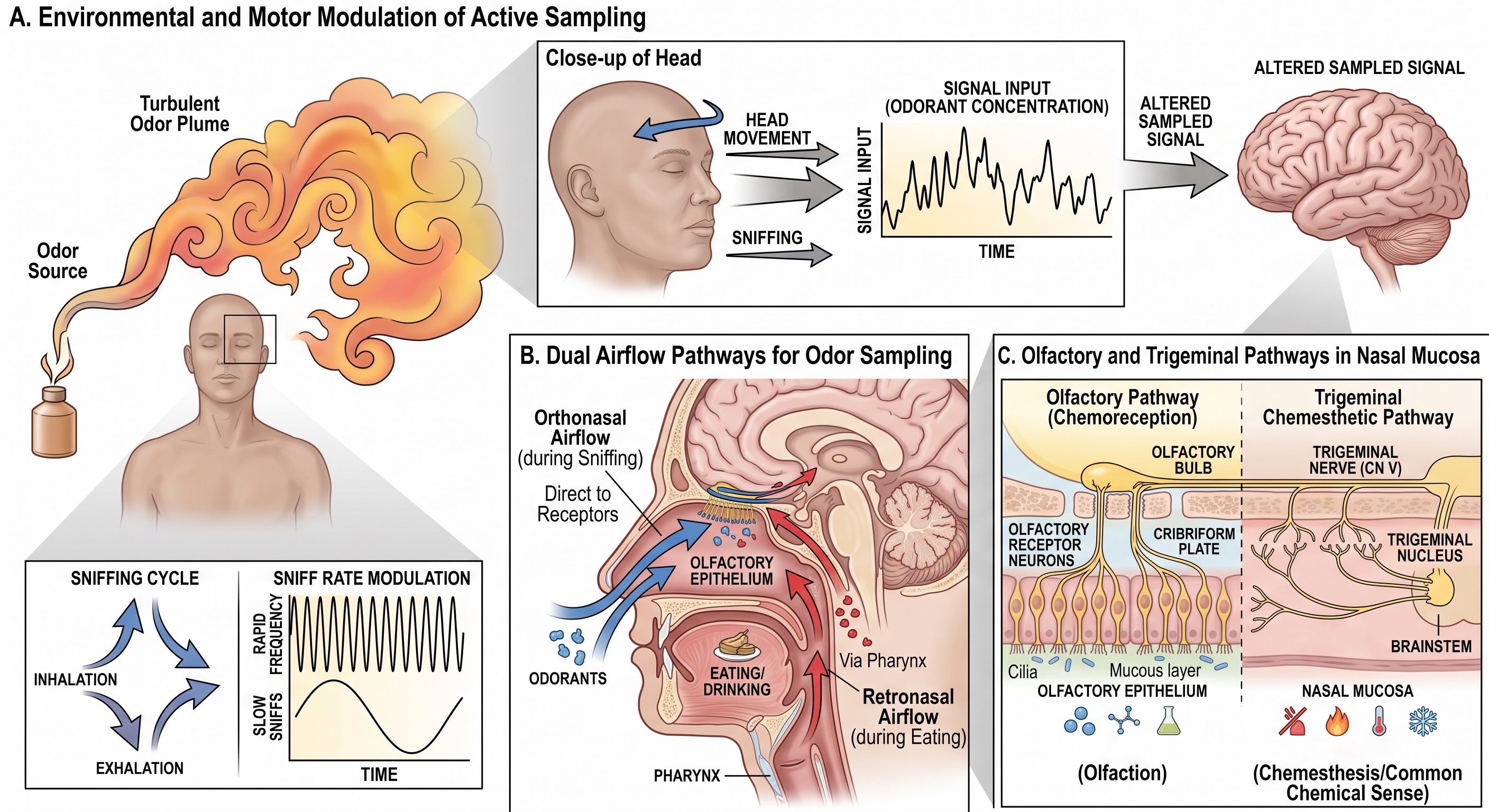
Odor is sampled through breathing
An odorant must reach the olfactory epithelium in the gas phase. Whether it does so depends not only on the molecule but also on temperature, vapor pressure, solubility, adsorption to nearby surfaces, and the airflow between source and observer. Once released, odorants are carried mainly by moving air. Turbulence breaks an odor plume into filaments and gaps, so a source may produce intermittent encounters rather than a smooth gradient that simply grows stronger with proximity.
This physics limits spatial precision, but it does not make odor direction unknowable. Humans can follow a scent trail across the ground, improve with practice, and perform worse when information from the two nostrils is artificially made identical [@porter2007tracking]. Other species may add larger nostril separations, mobile noses, rapid head movements, and specialized locomotor search strategies. Odor localization is therefore an active inference built from changing concentration, bilateral differences, wind direction, movement, and prior sampling rather than from a fixed receptor sheet that directly maps external space.
Sniffing is part of this active sensing. An inhalation transports molecules toward the olfactory epithelium and divides the chemical stream into respiratory samples. Changing sniff depth or duration changes which molecules reach which epithelial regions and when they arrive. Head movement and locomotion alter the plume itself from the animal’s point of view. As with eye movements in vision and palpation in touch, the motor act that acquires the signal helps determine the signal that the nervous system receives.
Odorants reach the epithelium by two routes. In orthonasal olfaction, molecules enter through the nostrils from the external environment. In retronasal olfaction, volatile molecules released in the mouth travel upward through the nasopharynx during chewing and exhalation. The receptor organ is the same, but the route of access and behavioral context differ. Orthonasal smell commonly identifies something before it enters the mouth; retronasal smell helps identify what is already being eaten.
Not every chemical sensation in the nose is olfactory. Free endings of the trigeminal system respond to irritants, cooling agents, and pungent chemicals. Ammonia can sting, menthol can feel cool, and high concentrations of many odorants can burn even when their olfactory qualities are weak. A single substance may activate both olfactory and trigeminal pathways. This nasal chemesthesis belongs to the somatosensory and pain systems discussed earlier, not to the olfactory nerve itself.
Olfactory sensory neurons transduce airborne chemicals
The olfactory epithelium is a sensory organ
The main olfactory epithelium occupies specialized mucosa high in the nasal cavity, along portions of the superior septum and upper nasal conchae. Its surface is covered by mucus produced by local glands and epithelial cells. Odorants must enter this aqueous layer before reaching receptor proteins in the sensory membrane.
The principal receptor cell is a bipolar olfactory sensory neuron. A short dendrite ends in a knob near the epithelial surface, from which several long, nonmotile cilia extend into the mucus. These cilia carry the molecular machinery for odor transduction. At the opposite pole, a thin unmyelinated axon joins neighboring axons into small bundles, passes through openings in the cribriform plate, and enters the olfactory bulb. Collectively, these axons constitute cranial nerve I.
Several non-neuronal populations maintain the receptor organ. Sustentacular cells support epithelial metabolism, ionic balance, and barrier function. Globose basal cells supply much of the continuing replacement of epithelial cells under ordinary conditions, while normally quiescent horizontal basal cells can be recruited after more substantial injury. Mucus and odorant-processing enzymes help control access to the cilia. The epithelium is therefore a renewing tissue, not merely a sheet of exposed nerve endings.
This arrangement differs sharply from taste. In a taste bud, an epithelial receptor cell transduces the stimulus and communicates with a separate first-order cranial neuron. In olfaction, the receptor cell is itself the first-order neuron. It generates action potentials and sends its own axon into the central nervous system. Somatosensory receptors provide a second comparison: many are specialized endings of pseudounipolar neurons whose somata lie in sensory ganglia, whereas olfactory-neuron somata remain within the receptor epithelium.
Olfactory sensory neurons can be replaced during adult life, but this fact should not be turned into a claim of effortless regeneration. A newly generated neuron must mature, select a receptor program, extend an axon through the cribriform plate, and join an appropriate bulbar target. Severe epithelial damage, disrupted basal-cell populations, scarring, inflammation, or central injury can leave recovery incomplete or distorted.
A cAMP cascade converts binding into spikes
Most familiar airborne odorants are detected by members of the large canonical odorant-receptor family first identified by Linda Buck and Richard Axel [@buck1991novel]. These receptors are G-protein-coupled receptors concentrated in the ciliary membrane. Binding begins a well-established transduction cascade:
- An odorant activates an odorant receptor.
- The receptor activates the olfactory G protein Golf [@jones1989golf].
- Golf activates type III adenylyl cyclase, increasing intracellular cAMP [@bakalyar1990adenylyl].
- cAMP opens cyclic-nucleotide-gated cation channels, allowing sodium and calcium to enter [@nakamura1987cyclic].
- Calcium opens a calcium-activated chloride conductance that includes ANO2. Because olfactory cilia maintain an unusually high internal chloride concentration, chloride leaves the cell and produces further depolarization [@stephan2009ano2].
- If the receptor potential is sufficient, the neuron generates action potentials that travel to the olfactory bulb.
The chloride current is often described as amplification, and it can contribute strongly to the receptor potential. Its effect on spike output and behavior is nevertheless more complicated than a one-way gain control: deleting ANO2 alters sensitivity, timing, sparsity, and tracking differently across experimental conditions. The safe generalization is that the cation and chloride currents jointly shape the conversion from receptor binding to spikes.
The cascade also adapts. Calcium–calmodulin interactions reduce the sensitivity of cyclic-nucleotide-gated channels, receptor and second-messenger mechanisms change during continuing stimulation, and central responses decline when an odor is repeated. The fading of a room odor after several minutes therefore has several possible contributors. Peripheral adaptation begins in the receptor neuron, but habituation and expectation continue to alter activity in bulb and cortex.
odorant → receptor GPCR → Golf → adenylyl cyclase III → cAMP → cation and chloride currents → action potentials
Hundreds of receptors create a combinatorial code
One receptor identity per neuron—usually
The human genome contains roughly 400 intact canonical odorant-receptor genes, together with several hundred related sequences that have become pseudogenes or truncated genes [@niimura2003human; @olender2020annotation]. The number is large compared with the receptor families used for taste, but it is tiny compared with the number of distinguishable odor experiences. Olfaction therefore cannot assign one receptor to coffee, another to rose, and another to smoke.
A mature canonical olfactory sensory neuron generally expresses one odorant-receptor gene from one allele [@chess1994allelic]. Neurons expressing a common receptor identity are distributed across an epithelial territory rather than packed into one compact receptor organ. The rule is biologically powerful but not exceptionless. Specialized olfactory-neuron classes use other receptor families, and modern single-cell measurements reveal occasional departures from a perfectly exclusive one-gene state. For the canonical pathway, however, receptor identity remains the principal molecular label carried by the neuron toward the bulb.
Receptor tuning is many-to-many. One receptor can respond to several structurally related or even rather different molecules, and one odorant can activate several receptor types. The relative activity across the receptor population forms a combinatorial code [@malnic1999combinatorial]. Changing concentration changes that pattern: additional receptors may be recruited, relative response strengths may shift, and a molecule can acquire a different perceived quality at high concentration.
Natural mixtures add another complication. Receptor responses are not always the linear sum of their components. Molecules can compete for a binding site, one component can antagonize activation produced by another, and the airflow and mucus can favor some molecules over others. Receptor-level antagonism can help prevent population activity from growing without bound as mixtures become more complex [@reddy2018antagonism]. The peripheral code is therefore a weighted and dynamic population response, not a binary list of all molecules present.
Genetic variation makes the code partly individual. Variants of OR7D4 alter the perceived intensity and pleasantness of androstenone, which some people describe as unpleasant, others as sweeter, and others barely smell at all [@keller2007variation]. A larger receptor survey found functionally important polymorphisms across much of the sampled human receptor repertoire; two individuals were estimated to differ functionally at more than 30% of their receptor alleles on average [@mainland2014missense]. Culture and learning matter greatly, but people also begin with somewhat different peripheral sensors.
The epithelium is distributed, but not random
The traditional account divided the mouse olfactory epithelium into several broad zones. Within each zone, neurons choosing a particular receptor were often portrayed as scattered almost randomly. That description captured the absence of compact receptor patches, but it understated the spatial order.
Two large mouse studies published in 2026 mapped nearly the full receptor repertoire across the epithelium and bulb. They found that receptor populations occupy reproducible average positions along continuous epithelial gradients and overlapping stripes. The epithelial arrangement is aligned with the locations to which those neurons project in the olfactory bulb [@brann2026spatial; @bintu2026spatial]. One study also related the map to developmental molecular gradients; the other identified two epithelial axes mirrored along corresponding bulbar axes and found spatially differentiated responses to several social and predator odors.
These results update the textbook model in an important but bounded way. Neurons carrying one receptor identity are still dispersed over territory rather than collected into a single receptor spot. The new finding is that their distribution is spatially biased and stereotyped, not arbitrary. The studies were performed in mice. Whether the human epithelium contains the same fine-grained organization, and what computation the arrangement supports in different species, remain open questions.
Chemical responsiveness is evolutionarily ancient and predates nervous systems, but the vertebrate olfactory epithelium, receptor families, bulb, and cortical circuits are highly derived biological structures. Modern smell is not simply bacterial chemotaxis enlarged inside a skull.
Comparative analysis of more than 1,500 vertebrate genomes shows that chemoreceptor families have undergone repeated lineage-specific expansions and losses. Repertoire size covaries with habitat, diet, olfactory-organ morphology, and other ecological variables, but no single evolutionary exchange explains all of the variation [@policarpo2024diversity]. Marine transitions, for example, have repeatedly reduced several chemoreceptor families, whereas some terrestrial lineages have expanded them.
Humans possess many olfactory-receptor pseudogenes and fewer intact canonical receptors than several other mammals. That history should not be reduced to a simple bargain in which vision replaced smell. Nor can receptor-gene count be read as a direct scale of overall olfactory performance. Receptor diversity, epithelial area, copy number per receptor type, airflow, bulb circuitry, learning, and the behavioral question being asked all contribute.
The often repeated estimate of 10,000 odors has no secure experimental derivation. A 2014 psychophysical study extrapolated from discrimination among selected mixtures and proposed that humans could distinguish at least one trillion olfactory stimuli [@bushdid2014trillion]. Subsequent analyses showed that the result depends strongly on how mixture similarity, sampling, and perceptual-space volume are defined [@gerkin2015trillion; @meister2015trillion].
There is presently no defensible single count. Receptor combinations do not translate directly into equally spaced percepts, and mixtures that differ chemically can smell similar while closely related mixtures can sometimes be distinguished. The important conclusion is not a spectacular number but the high dimensionality and flexibility of human odor discrimination.
The olfactory bulb is smell’s first sensory processor
Axons from olfactory sensory neurons enter the ventral surface of the bulb and terminate within spherical neuropil structures called glomeruli. In the best-studied mammalian systems, sensory neurons expressing a common receptor identity converge onto a small, stereotyped set of glomeruli in each bulb [@mombaerts1996visualizing]. The distributed epithelial population is thereby reorganized into a bulbar map whose glomeruli retain information about receptor identity.
Each glomerulus is more than a bundle of incoming axons. Olfactory sensory neurons contact the apical dendrites of mitral cells and tufted cells, the principal bulbar output neurons, as well as local interneurons. Periglomerular circuits shape activity near the glomerular layer. Deeper granule cells form reciprocal dendrodendritic interactions with mitral and tufted cells. These inhibitory networks influence gain, timing, synchrony, and the separation or coordination of output patterns. Describing all of them as simple lateral inhibition is convenient but incomplete.
Convergence provides a strong organizing step. Many sensory neurons carrying the same receptor identity can contribute to one glomerular channel, while local circuits compare that channel with activity elsewhere in the bulb. At the same time, odor information is represented in temporal structure. Bulbar responses unfold within a respiratory cycle, and different mitral and tufted cells can respond at different phases and latencies. A glomerular map is therefore not a static image stamped onto the bulb; it is a spatially organized, time-varying population response.
The bulb also receives extensive descending input. Piriform cortex and the anterior olfactory nucleus project back to bulbar interneurons and output cells, while cholinergic, noradrenergic, serotonergic, and other systems signal behavioral state. In awake mice, suppressing piriform feedback increases the similarity of mitral-cell responses, indicating that descending activity can help decorrelate bulbar output [@otazu2015feedback]. The first central processor is already recurrent, in keeping with the general sensory architecture developed in Chapter 21.
The first route is mainly ipsilateral
The crossing logic differs from the pathways emphasized in somatosensation. Receptor neurons in one nasal epithelium project to the bulb on the same side, and bulbar output reaches ipsilateral primary olfactory areas first. There is no obligatory decussation comparable to the spinal or medullary crossings of the major somatosensory systems.
The two sides do communicate. The anterior olfactory nuclei exchange information through the anterior commissure, and bilateral cortical and subcortical circuits combine the two streams. Human intracranial recordings provide a useful temporal demonstration. Odor identity delivered to the nostril ipsilateral to a recorded piriform cortex was represented about 480 milliseconds earlier, on average, than identity delivered to the contralateral nostril. During bilateral smelling, ipsilateral and contralateral information appeared in two separable epochs within one sampling episode [@dikecligil2023nostrils].
The system is therefore predominantly ipsilateral at its entrance, not permanently unilateral. Bilateral comparison can contribute to tracking and localization, while central interactions allow the two nostrils to inform a common percept.
From glomerular channels to cortical ensembles
Primary olfactory cortex comes before the thalamus
Olfaction is commonly said to “bypass the thalamus.” That phrase is useful only if its scope is made explicit. Bulbar output can reach primary olfactory cortex without first passing through an obligatory thalamic relay. Olfaction therefore differs from the canonical receptor → first-order thalamus → primary neocortex sequence. It does not follow that the thalamus is absent from olfactory perception.
Piriform cortex communicates with orbitofrontal cortex by both direct corticocortical routes and an indirect route through the mediodorsal thalamus. Human fMRI connectivity analyses found that attending to odor selectively strengthened forward coupling along the piriform → mediodorsal thalamus → orbitofrontal pathway [@plailly2008attention].
The distinction fits the thalamic framework in Section 21.3. Olfaction lacks an obligatory thalamic relay before primary olfactory cortex, but later olfactory processing recruits thalamic circuits that regulate cortical communication, attention, comparison, and report. The mediodorsal nucleus is not simply the delayed equivalent of the visual LGN or somatosensory VPL/VPM. It participates at a later stage in a pathway that has already reached cortex.
Nor should the direct bulbar route be relabeled a unitary “limbic shortcut.” Bulbar output is distributed across several olfactory cortical and amygdalar targets, and those regions interact with orbitofrontal, insular, striatal, hypothalamic, and medial temporal networks. In rodents, direct bulbar projections to lateral entorhinal cortex are well established. Direct bulb-to-entorhinal projections have not yet been anatomically confirmed in humans, and human single-neuron data suggest that entorhinal odor coding emerges later than activity in piriform cortex and amygdala [@kehl2024single]. Short anatomical routes do not place smell outside ordinary recurrent and multisynaptic brain processing.
Odor identity is not odor value
An odor representation can carry several partially separable kinds of information:
- identity—what source or substance the odor is taken to indicate;
- intensity—how strong the current signal is;
- quality—the perceptual character used to compare it with other odors;
- value—how attractive, aversive, or important it is in the current state;
- meaning—what the odor predicts from prior experience.
These variables interact but are not interchangeable. Concentration can alter both quality and pleasantness. A faint indole concentration may contribute to a floral mixture, whereas a stronger concentration can smell fecal. A food odor can become less attractive during a meal, and an initially neutral odor can become aversive after it predicts nausea. The receptor pattern constrains the percept, but current value is constructed from that pattern together with learning, context, and physiological state.
This distinction parallels the taste chapter. Sweetness and palatability are not the same variable; neither are odor identity and odor value. Some nonhuman species possess relatively selective receptor and circuit channels for ecologically urgent chemicals, and humans may share initial biases toward or away from some odor classes. But human odor value cannot be reduced to a universal receptor-level axis running from perfume to decay. Familiarity, naming, concentration, culture, recent experience, and bodily need can all move the same odor along the evaluative dimension.
Piriform cortex, amygdala, and orbitofrontal cortex contribute different but overlapping information. Human single-neuron recordings found that piriform neurons more strongly represented chemical identity, while amygdala neurons tracked participants’ own pleasantness judgments [@kehl2024single]. The odor set in that study was weighted toward pleasant examples, and intensity can interact with valence, so the finding should not be turned into a complete cellular map of liking and disliking. Orbitofrontal activity is especially sensitive to reward value, expectation, and changing motivational state, but it is embedded in reciprocal interactions rather than serving as a final meter attached to an otherwise finished percept.
The broader principle is that olfactory processing does not progress from objective identity to an optional emotional label appended at the end. Value-related and identity-related activity interact at several stages, including feedback to bulb and piriform cortex. Yet preserving the conceptual distinction remains essential: recognizing coffee, judging it strong, liking it this morning, and using it as a cue that breakfast is ready are different accomplishments.
Disgust is more than sensory aversion
Taste and olfaction are unusually potent routes into disgust. Taste evaluates chemicals that have reached the mouth; olfaction can warn of decay, excreta, or contamination before contact. Yet disgust is not a taste quality, an odor quality, or merely the unpleasant end of a value scale. An odor can be unpleasant because it is intense or irritating without being disgusting, and an aversive taste can elicit oral rejection without requiring the full emotional state ordinarily meant by disgust.
Distaste, nausea, and disgust are not synonyms
The distinctions developed in Section 28.4 are useful here. Distaste, in the narrow physiological sense, is a patterned oral rejection response: gaping, tongue movements, spitting, interruption of ingestion, and related actions. In rats, characteristic ingestive and rejection patterns survive decerebration, demonstrating that caudal brainstem circuits can organize them without the forebrain machinery needed for ordinary conscious appraisal [@grill1978decerebrate]. Nausea is a state of visceral malaise, and vomiting is a centrally patterned motor act. Disgust is broader: it combines interpretation of what is present, bodily feeling, facial and attentional changes, memory, and tendencies to withdraw, reject, avoid, or cleanse.
These processes frequently cooperate, but none is a synonym for the others. A bitter liquid can be rejected without later nausea. Nausea can occur without vomiting. A contaminated object seen across a room can evoke disgust without any chemical entering the nose or mouth. Conversely, people can judge an odor unpleasant without construing its source as contaminating. Chemical sensation supplies especially powerful evidence about possible ingestion and bodily harm, but the resulting state depends on the source attributed to the signal, what has been learned about it, and what actions remain possible.
The facial expression of disgust may itself alter sampling. When participants posed fear and disgust expressions, the two configurations had opposite effects: fear increased nasal volume and inspiratory airflow, whereas disgust reduced them [@susskind2008fear]. That result is consistent with limiting further exposure at the nose and mouth. It is not proof that every disgust expression evolved for pathogen avoidance, but it provides a concrete link between an emotional action pattern and control of sensory input.
The insula is involved, but it is not a disgust center
A famous early social-neuroscience experiment helped make disgust an insula story. Participants inhaled foul odorants and, in a separate condition, watched video clips of people expressing disgust. Both conditions produced overlapping activity in anterior insula and, less strongly, anterior cingulate cortex. The authors proposed that observing another person’s disgust reactivated the neural representation used to experience disgust oneself [@wicker2003insula].
The basic observation has held up well. Viewing disgusted faces reliably engages anterior insula and amygdala, and experiencing disgust directly — through foul odors or bitter tastes — engages overlapping insular territory. Neuroimaging of disgust is not a case of a finding that failed to replicate. What did not survive was the interpretation built on top of it.
Two problems recur. The first is that activation is observational. A region that becomes active during a task need not be necessary for it, and need not be doing the thing the experimenter names. Anterior insula is among the most frequently activated regions in the entire imaging literature: it responds during pain, uncertainty, effort, risk, interoception, error detection, and attention to bodily state. Finding it active during disgust constrains interpretation much less than it first appears, because finding it active is close to the default.
The second problem is specificity. If anterior insula were the disgust region and amygdala the fear region, each should respond selectively to its own emotion. Neither does. Amygdala responds to disgusted faces as well as fearful ones, and more generally to faces that are arousing, ambiguous, or biologically significant. Insula responds during fear and anger as well as disgust. The tidy mapping of one structure to one emotion was never what the data showed; it was a simplification imposed on results that were considerably messier.
Intervention evidence points the same way. Direct stimulation of the left insula in surgical patients produced a broad decrement in facial-emotion recognition, significant for disgust but not cleanly category-specific [@papagno2016disgust]. Patients who had undergone unilateral insular resection showed some general impairment in emotion recognition but no selective deficit for disgust [@boucher2015insula]. Unilateral lesions and postoperative reorganization limit what follows from preserved performance, so these are not decisive. Taken with the imaging results, though, they fit a network account rather than a module.
A defensible positive claim survives all of this. Anterior insula is where visceral, gustatory, and olfactory signals converge and are re-represented in a form available to conscious feeling. Disgust engages it strongly because disgust is unusually visceral among the emotions, not because the region is dedicated to disgust. That account predicts the overlap Wicker and colleagues observed, predicts that lesions produce graded rather than categorical deficits, and predicts that other emotions with strong bodily components will recruit the same territory. It asks the region to do something more general than house an emotion, and something more specific than participate in everything.
Disgust also extends well beyond food and contamination. People describe betrayal, exploitation, and various sexual or religious violations as disgusting, and disgust sensitivity correlates with the severity of moral judgments, most strongly for violations framed around purity or sanctity [@donner2023specificity]. Whether incidental disgust causally makes moral judgment harsher is much less clear; the experimental literature on that point is weak and has not replicated well [@landy2015disgust; @ghelfi2020gustatory]. What survives is partial continuity and convergence between chemical rejection and social judgment, not one neural module carried unchanged from spoiled food to morality.
Olfaction, memory, and learned meaning
Odors often appear to summon memories with unusual immediacy. Proust’s description of a madeleine releasing a long-unavailable childhood scene gave the phenomenon its familiar literary name. The anatomy makes the association plausible: piriform and amygdalar olfactory regions interact with entorhinal cortex, hippocampus, orbitofrontal cortex, and other structures involved in learning. But “plausible” should not be converted into a unique direct-access memory system.
Cross-modal experiments provide a more precise result. Odors can evoke autobiographical memories that participants rate as more emotional and more evocative than memories cued by corresponding pictures or sounds. In the same experiments, odor-cued memories were not consistently more vivid or more specific [@herz2004naturalistic]. In associative-memory studies, odors produced recall accuracy comparable to other cues while increasing the emotional response to what was recalled [@herz1998odors]. The Proust effect is therefore best described as an effect on emotional reliving under some conditions, not as proof that smell stores more accurate memories or bypasses ordinary retrieval.
Prior experience is central. An odor acquires meaning by occurring with a food, room, person, illness, season, occupation, or danger. The same molecule can therefore evoke very different autobiographical and evaluative responses across individuals. Odor naming is often difficult without context, yet a verbal or visual cue can rapidly stabilize identification. The cross-modal and semantic responses found in human piriform and amygdala neurons fit this learned organization [@kehl2024single].
Olfaction is thus strongly connected to memory because odors become components of events and predictions. It is not “the memory sense” in isolation from the rest of the brain. Music, faces, places, and words can also be powerful cues, and their relative effectiveness depends on what was encoded, how distinctive the cue is, and what the task asks a person to recover.
When smell is lost or distorted
Olfactory disorders can reduce the amount of information available or alter the quality of what is perceived. The principal terms distinguish these possibilities:
- anosmia is an absence of measurable smell;
- hyposmia is reduced smell;
- parosmia is a distorted perception produced by an odor that is present;
- phantosmia is an odor percept without a corresponding external odorant.
The pathway suggests several sites of failure. A conductive disorder prevents molecules from reaching the olfactory epithelium because of swelling, polyps, obstruction, or altered airflow. An epithelial disorder damages receptor neurons, supporting cells, mucus, or local inflammatory control. Head trauma can tear the delicate axon bundles where they pass through the cribriform plate or damage the bulb and orbitofrontal surface. Tumors, surgery, seizures, and other central disorders can alter bulbar or cortical processing. A complaint that food no longer “tastes right” must therefore be separated into basic taste, orthonasal smell, retronasal smell, oral somatosensation, and flavor integration, as emphasized in Chapter 28.
COVID-19 supplied an unusually visible example of epithelial olfactory dysfunction. Early molecular studies found the major SARS-CoV-2 entry machinery concentrated primarily in sustentacular, basal, and vascular-associated cells rather than mature olfactory sensory neurons [@brann2020nonneuronal]. Work in human tissue and hamsters later found widespread downregulation of odorant-receptor and signaling genes in sensory neurons, accompanied by altered nuclear organization, even though the neurons themselves were rarely infected [@zazhytska2022nuclear]. COVID-related anosmia should therefore not be summarized simply as a virus killing smell neurons. Supporting-cell injury, inflammation, ciliary disruption, altered gene expression, and subsequent repair can all contribute.
Parosmia often appears during recovery from postviral or traumatic loss. Familiar foods may acquire burnt, sewage-like, chemical, or otherwise alien qualities. Gas-chromatography–olfactometry studies have isolated a small set of highly odor-active molecules that commonly trigger distortions in affected participants [@parker2022parosmia]. This identifies useful stimulus regularities but does not settle the neural mechanism. Selective survival or recovery of receptor populations, altered receptor ratios, abnormal axonal reconnection, and changed central interpretation remain plausible and potentially interacting explanations.
Olfactory training repeatedly exposes a person to a set of odors and is often used during recovery. Its efficacy is difficult to evaluate because etiology, disease duration, spontaneous recovery, adherence, and outcome measures differ across studies. In a 2025 placebo-controlled trial of participants with long-standing post-COVID dysfunction, twelve weeks of conventional odor training did not improve objective olfactory scores more than odor-free placebo training; parosmia was associated with poorer response [@mogensen2025training]. That result does not prove that training is ineffective for every cause or at every stage, but it cautions against presenting one protocol as a uniformly established restoration treatment.
The consequences of olfactory loss extend beyond pleasure. Smell contributes to detecting smoke, gas leaks, spoiled food, and changes in the home environment. It supports appetite, food choice, and the sense of familiarity attached to people and places. Distortion can be more disabling than simple reduction because commonly eaten foods become repellent. These effects arise because olfaction normally participates in safety, nutrition, social recognition, and learned environmental prediction—not because it is an optional aesthetic sense.
Flavor revisited
Section 28.1 distinguished taste from flavor. The olfactory side of that distinction can now be made anatomically explicit. During chewing, volatile molecules released from food move retronasally toward the same olfactory epithelium used in orthonasal smell. Olfactory sensory neurons do not know whether a molecule arrived from a cup in front of the face or from food in the mouth. The routes nevertheless differ in airflow, timing, accompanying movements, and the gustatory and somatosensory signals with which the odor occurs.
Retronasal olfaction (Figure 29.1) supplies much of the information that distinguishes one food from another. Blocking nasal airflow can leave sweetness, saltiness, sourness, bitterness, umami, temperature, texture, and chemesthetic burn relatively intact while removing much of a food’s recognizable identity. When airflow is restored during chewing, the returning odor is commonly experienced as belonging to the food in the mouth rather than as a separate event in the nose. This oral referral is one reason people describe olfactory loss as loss of taste.
Flavor is not reducible to smell. It combines gustation, retronasal olfaction, oral touch, viscosity, temperature, chemesthesis, expectation, and learned consequences. The relative contribution of each signal changes with the food and task. Retronasal odor often supplies food-specific identity; taste contributes a smaller set of chemical qualities with strong ingestive significance; somatosensation supplies texture and temperature; and all of them can alter value and action.
Human imaging now provides direct evidence for learned taste–odor integration. After participants learned particular taste and retronasal-odor pairings, the paired signals produced partially shared, flavor-specific activity patterns in dysgranular and agranular insular cortex, especially in more ventral anterior regions [@khorisantono2025flavour]. The result supports convergence of associated taste and odor information. It does not establish one exclusive cortical “flavor center,” nor does the involvement of insula make taste, pain, interoception, and flavor the same function; Section 29.8 developed the same caution for disgust. The more defensible conclusion is that neighboring and interacting insular territories combine oral and bodily signals at several levels of processing.
Looking ahead: from sensing to valuation and action
Olfaction completes the chemical-sense sequence begun in the taste chapter. Molecules carried through air are actively sampled by breathing; receptor neurons convert binding into spikes; receptor-defined populations converge in the bulb; bulbar channels are transformed into distributed cortical ensembles; and later thalamic, orbitofrontal, amygdalar, insular, striatal, hypothalamic, and medial temporal interactions help determine identity, value, memory, and action.
The chapter also supplies a final variation on the unit’s shared plan. Olfaction has a receptor neuron but no separate peripheral ganglion, a first central processor but no obligatory early decussation, primary cortex before thalamus, and paleocortex rather than granular sensory neocortex. The system is distinctive without being exempt from population coding, convergence, recurrence, parallel pathways, and state-dependent control.
Unit V changes the level of emphasis rather than introducing action for the first time. A sniff is already a movement that controls sensory input; following a plume is already closed-loop behavior; and the value of a food odor is already changing the probability of approach. The next unit turns more directly toward the circuits that compare alternatives, assign current value, select among possible actions, and organize movement.
Reasonably settled:
- Olfactory sensory neurons are bipolar first-order neurons whose receptor-bearing cilia contact nasal mucus. Canonical odorant receptors usually signal through Golf, adenylyl cyclase III, cAMP, cyclic-nucleotide-gated channels, and a calcium-activated chloride conductance.
- Mature canonical sensory neurons generally express one principal odorant-receptor identity. Odorants are encoded through combinatorial population activity rather than one receptor per perceived odor.
- Neurons sharing a receptor identity converge onto stereotyped glomerular targets in the mammalian olfactory bulb. Mitral and tufted cells distribute the processed output through parallel pathways, and the bulb receives strong cortical and neuromodulatory feedback.
- The initial pathway is predominantly ipsilateral. Bilateral information is combined through commissural and later central interactions rather than through an obligatory early decussation.
- Piriform cortex receives direct bulbar input and uses a distributed population code that does not preserve a simple point-to-point copy of the glomerular map.
- Primary olfactory cortex is reached without an obligatory preceding thalamic relay. Mediodorsal thalamus nevertheless contributes to later communication with orbitofrontal cortex, especially during attention and identification.
- Odor identity, intensity, value, and learned meaning are related but separable. Retronasal olfaction makes a major contribution to food identity and interacts with taste and oral somatosensation to construct flavor.
- Taste and olfaction can powerfully trigger disgust, but disgust is not a dedicated chemical-sense channel or an exclusive function of anterior insula.
Still being refined:
- How complex mixtures are encoded. Receptor agonism, antagonism, airflow, concentration, adaptation, bulbar circuits, and cortical learning all contribute, but no complete account predicts mixture perception from chemistry alone.
- How stable identity is extracted across concentration and context. The same source can generate very different peripheral patterns as distance, airflow, sniffing, and concentration change.
- How general the newly described epithelial map is. The detailed receptor gradients and stripes reported in 2026 were demonstrated in mice; their fine organization and function in humans and other species remain unknown.
- What local piriform organization computes. New mouse evidence indicates that nearby neurons share somewhat more bulbar input and odor tuning, but this is not yet a canonical chemotopic map.
- How much odor value is inherited and how much is learned. Selective innate channels exist in several nonhuman species, but human pleasantness and aversion are strongly altered by concentration, context, culture, experience, and bodily state.
- Why parosmia occurs and why recovery varies. Peripheral receptor loss and regrowth, axonal targeting, epithelial inflammation, and central reinterpretation may all contribute.
- What is distinctive about odor-cued autobiographical memory. Emotional evocation has empirical support; superior accuracy, vividness, or a unique direct-access memory mechanism does not.
- How much mechanism is shared across oral distaste, nausea, core disgust, observed disgust, and moral condemnation. Some components overlap, but evidence for a single insular system or a general causal effect of incidental disgust on moral judgment is inconsistent and strongly moderated by method, target, culture, and moral domain.
- Whether humans possess behaviorally important pheromones in the strict biological sense. Social odors can influence perception and physiology, but no generally accepted human pheromone system has been established.