19 Modulating the neuron’s state
Neuromodulation, Volume Transmission, and the Systems That Set the Brain’s State
Chapter 18 explained fast electrical signaling from first principles while deferring two related topics. One was how synapses change their strength with use—the cellular basis of memory, taken up in Chapter 20, the final chapter of this unit. The other was the metabotropic receptor, introduced through the image of a doorbell rather than the locked gate of an ionotropic receptor. When a signaling molecule binds an ionotropic receptor, the receptor’s pore opens directly. Binding to a metabotropic receptor instead initiates a biochemical cascade inside the neuron. This is the slower chemistry of neural signaling: rather than carrying the moment-to-moment content of a message, it often changes the conditions under which messages are generated, received, and modified. Some cascades ultimately open or close ion channels; others alter enzymes, synapses, or gene expression.
The unit’s organizing claim has been that neural signaling operates across many timescales, and that fast electrical signaling is the most evolutionarily recent—and the most metabolically expensive—layer. The fast layer carries information from moment to moment. This chapter examines a slower layer that regulates how that information is carried. Across populations of neurons, neuromodulatory systems can shift arousal, alter excitability, and change the conditions under which synapses undergo plasticity. Their consequence is something fast point-to-point signaling cannot produce on its own: a coordinated brain state. Metabotropic receptors provide much of the cellular machinery for these changes, while widely branching projections and more diffuse forms of release give many modulatory systems their anatomical reach.
We will begin with the intracellular machinery of metabotropic receptors. We will then distinguish point-to-point synaptic transmission from signaling that spreads more broadly through the surrounding tissue, and examine why neurotransmitter and neuromodulator describe two kinds of job rather than two fixed classes of molecule. The chapter next turns to the small, deep nuclei whose widely branching projections influence the operating state of large portions of the brain, and to the anatomical arrangements that allow relatively few neurons to affect so many targets. We will then consider how a striking number of prescribed and illicit drugs act upon this layer of signaling. The chapter closes with a question: why is the standard textbook list of neuromodulators narrower than the biology itself?
19.1 From the doorbell, downstream
Recall the architectural analogies for three membrane mechanisms depicted in Figure 18.2. An ionotropic receptor was a locked gate: neurotransmitter binding opens the pore directly, ions cross the membrane, and the effect begins within milliseconds. A voltage-gated channel was an emergency door that opens in response to the membrane’s own electrical state. A metabotropic receptor was a doorbell: a molecule binds outside the cell, but the receptor does not itself contain an ion channel. Instead, binding changes the receptor’s shape and engages molecular machinery on the inner surface of the membrane.
At the level needed here, most metabotropic neurotransmitter receptors belong to the family of G-protein-coupled receptors. When a signaling molecule binds, the receptor changes shape on its intracellular face and activates a G protein. Components of the activated G protein can act directly upon ion channels. They can also regulate enzymes that generate second messengers—small intracellular signals such as cyclic AMP, IP₃, and DAG. These molecules spread within the cell, activate protein kinases, and coordinate effects at targets some distance from the receptor. The extracellular signaling molecule is the first messenger; the intracellular signals it recruits are the second messengers.
A G-protein-coupled receptor, or GPCR, crosses the cell membrane but does not form a pore through it. When a signaling molecule binds outside the neuron, the receptor changes shape and activates a G protein attached to its inner surface. The name comes from the guanine nucleotides GDP and GTP, which act as a molecular switch. In its inactive state, the G protein holds GDP. Activation replaces GDP with GTP, allowing parts of the G protein to separate and regulate enzymes or ion channels. The G protein eventually converts GTP back to GDP, providing a built-in mechanism for ending the signal.
The letters attached to G proteins identify different families with different downstream effects. Gs commonly stimulates adenylyl cyclase and increases the second messenger cyclic AMP; Gi/o usually inhibits adenylyl cyclase and can also regulate potassium and calcium channels; Gq activates a pathway that releases calcium from stores inside the cell and activates protein kinase C. These are not rigid one-receptor–one-effect rules. The outcome also depends on the enzymes, channels, and other signaling machinery present in that particular neuron.
G proteins are only one way for a receptor to transmit a signal across the membrane. Some receptors contain their own enzyme activity, while others recruit enzymes attached to their intracellular surface. Steroid hormones take another route: because they can cross the membrane, they bind receptors inside the cell and can act more directly upon gene expression. The nervous system therefore uses several molecular solutions to the same general problem—allowing a signal outside a cell to alter what happens inside it.
Calcium can also serve as a second messenger. We encountered it in the last chapter when it entered the presynaptic terminal in the axon bouton and triggered neurotransmitter release. Chapter 20 will show calcium entering the postsynaptic spine through NMDA receptors and initiating synaptic plasticity. Metabotropic cascades can alter intracellular calcium in several ways, including indirectly regulating calcium channels in the membrane and releasing calcium from stores inside the cell.
Three consequences follow from this indirect route. Together, they explain why metabotropic signaling can alter the state of a neuron rather than merely produce a brief electrical current.
The first is amplification. An ionotropic receptor conducts current through its own pore. A metabotropic receptor, by contrast, can activate multiple G proteins; each G protein can regulate an enzyme that generates many second-messenger molecules, and each downstream enzyme can modify many target proteins. A small extracellular signal can therefore produce a much larger intracellular response. The fast synapse gains precision by keeping each effect local and bounded. The metabotropic cascade gains power through branching and multiplication.
The second is duration. An ionotropic current begins and ends rapidly as the neurotransmitter dissociates or is cleared and the channel closes. G-protein cascades involve a series of biochemical reactions, so their effects usually develop more slowly and can outlast the extracellular signal that initiated them. Depending on the pathway, those effects may persist for hundreds of milliseconds, seconds, minutes, or longer.
By changing whether ion channels open, how readily they open, or how many are present in the membrane, a metabotropic cascade can shift a neuron’s resting membrane potential. A neuron modestly depolarized at -60 mV rather than -70 mV is closer to the threshold for generating an action potential. At glutamatergic synapses, that depolarization also makes it easier to relieve the voltage-dependent magnesium block of NMDA receptors when glutamate arrives. A modulatory signal therefore need not produce the next message itself. It can change how the neuron responds to many subsequent messages.
The third consequence is breadth of action. A metabotropic cascade is not confined to opening or closing channels. It can alter enzymes, neurotransmitter release, receptor trafficking, the cytoskeleton, and—through signaling pathways that reach the nucleus—the transcription of genes. A brief signal at the membrane can therefore produce both immediate changes in excitability and much slower changes in the proteins the neuron manufactures. This is one bridge between signaling and lasting cellular change.
Ionotropic receptors can also initiate durable plasticity—the NMDA receptor is a central example—but they do so by recruiting downstream biochemical machinery. The brief ionic current begins the process; intracellular cascades carry it forward. Chapter 20 will examine how those cascades produce lasting changes in synaptic strength.
The relative slowness of metabotropic signaling is therefore not simply a limitation. It is a consequence of the same intermediate steps that permit amplification, persistence, and access to many cellular targets. Fast ionotropic signaling provides millisecond precision. Slower biochemical signaling changes the operating conditions under which that precision is used. Neurons require both, and ordinarily run both at once.
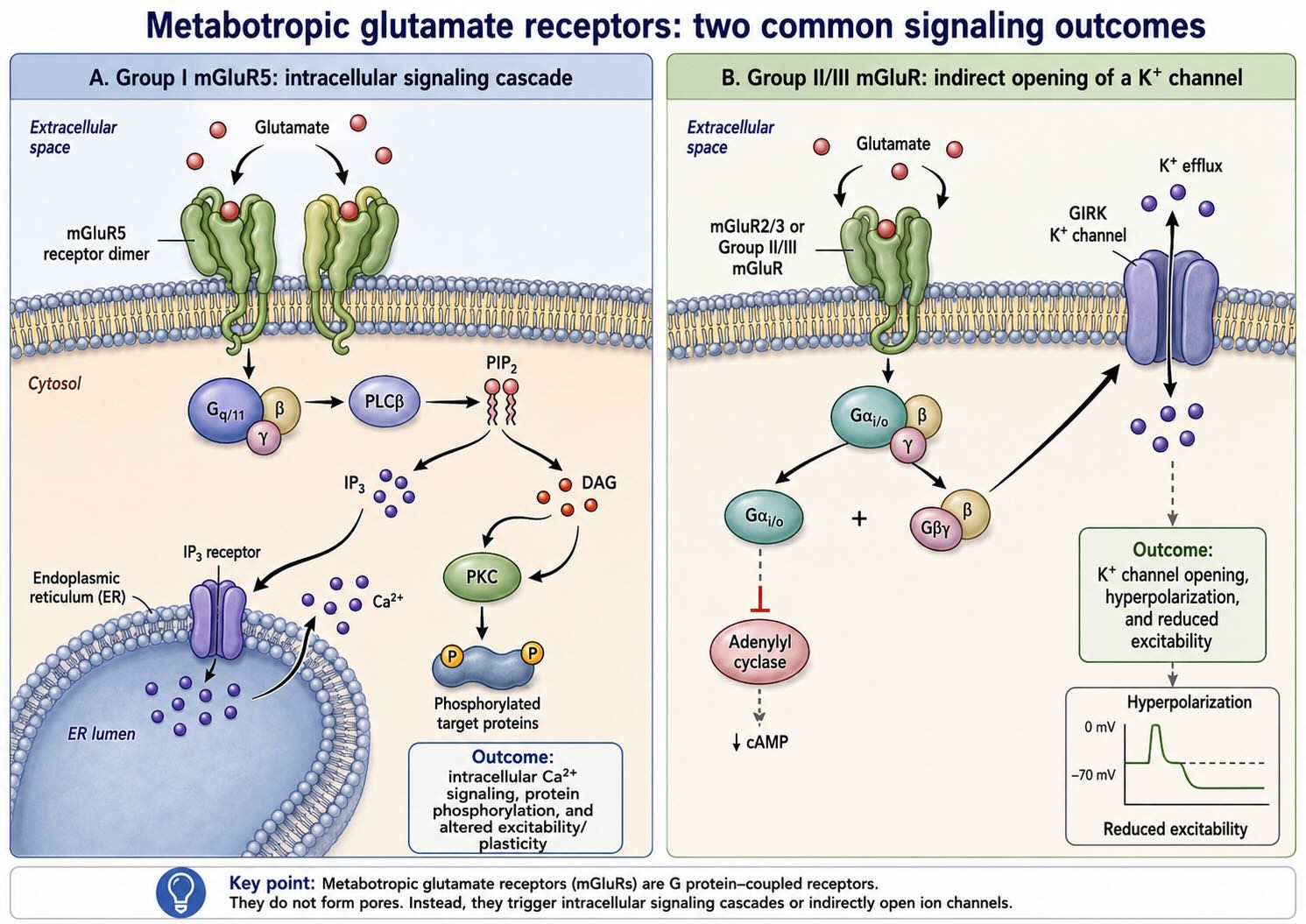
Glutamate provides a useful example because the same signaling molecule acts through both fast and slow receptors. At AMPA and NMDA receptors, glutamate opens an ion channel directly. AMPA receptors produce a rapid depolarizing current dominated by sodium entry. NMDA receptors also admit calcium when glutamate binding coincides with sufficient postsynaptic depolarization to relieve their magnesium block. The metabotropic glutamate receptors (mGluRs), by contrast, contain no pore. Glutamate binding activates G-protein pathways that alter ion channels, intracellular calcium, neurotransmitter release, and synaptic plasticity over longer timescales.
The mammalian nervous system expresses eight mGluR subtypes, organized into three groups according to their structure, pharmacology, and predominant G-protein coupling. Group I receptors—mGluR1 and mGluR5—are found mainly postsynaptically and generally couple to Gq proteins. They activate phospholipase C, mobilize calcium from intracellular stores, and activate protein kinase C. Their effects often increase neuronal excitability and support forms of synaptic plasticity.
Group II receptors—mGluR2 and mGluR3—and Group III receptors—mGluR4, mGluR6, mGluR7, and mGluR8—generally couple to Gi/Go pathways and reduce the activity of adenylyl cyclase. Many are positioned presynaptically, where they act as autoreceptors or heteroreceptors that limit neurotransmitter release and help regulate network activity. Individual subtypes also have additional locations and functions. The mGluR5 pathway illustrated in Figure 19.1 provides two examples of metabotropic glutamate pathways.
19.2 Two ways to send a message: wired and volume transmission
Having considered what happens when a signaling molecule reaches a receptor, we can turn to a different question: how does the molecule reach its target in the first place? Chemical signaling in the nervous system can be organized into two broad modes. The difference lies less in the identity of the molecule than in its addressing.
The first is the mode emphasized throughout the previous chapter. A presynaptic terminal lies immediately beside a defined region of postsynaptic membrane, separated from it by a narrow synaptic cleft. Transmitter released into the cleft crosses it within a fraction of a millisecond and encounters receptors concentrated on the postsynaptic cell. Transporters on nearby neurons and astrocytes, together with diffusion and, in some systems, enzymatic breakdown, limit how far and how long the signal spreads. This is wired transmission: a relatively private line between defined cellular partners. It is characteristic of rapid glutamatergic and GABAergic signaling, and its principal virtue is specificity. Activity in one neuron can influence a particular postsynaptic target within milliseconds.
The second mode is less tightly addressed. The releasing neuron may secrete its molecule from swellings along an axon or from other release sites that are not precisely aligned with a single postsynaptic membrane. The molecule enters the extracellular space—the fluid between cells—and diffuses through the surrounding tissue. Any cell within reach can respond if it expresses the appropriate receptor. This is volume transmission, named for the volume of tissue through which the signal spreads. Its principal virtue is reach: one neuron can influence many targets, including cells with which it forms no conventional synapse.
Volume transmission is locally analogous to hormonal signaling. A hormone enters the bloodstream and can be carried throughout the body, acting on cells that express the appropriate receptor. In volume transmission, a signaling molecule enters the brain’s extracellular fluid and acts on receptive cells within the surrounding tissue. The overview introduced paracrine signaling—the release of a molecule that diffuses to nearby cells—and noted that synaptic transmission can be understood as a tightly constrained form of it. Volume transmission allows that diffusion to extend beyond a single synaptic cleft. It remains local compared with endocrine signaling through the circulation, but it follows a similar addressing principle: receptor expression determines which cells respond.
These modes do not belong to two completely separate populations of neurons, and neither forms an absolute category. A neuron can make conventional synapses at some terminals and release transmitter more diffusely at others. Molecules released into a synaptic cleft can spill beyond it and activate extrasynaptic receptors, while signals usually associated with neuromodulation can also be released at anatomically defined synapses. Even so, the two modes have characteristic associations. Wired transmission often acts through fast ionotropic receptors clustered at a synapse. Volume transmission often acts through metabotropic receptors distributed across target cells, whose amplified and persistent effects are well suited to changing a cellular or circuit state rather than conveying a brief, precisely timed event.
| Wired transmission | Volume transmission | |
|---|---|---|
| Addressing | Point-to-point: defined presynaptic and postsynaptic partners | Diffuse: released into extracellular space |
| Spatial reach | Mainly across a single synaptic cleft | Through a local volume of tissue, potentially reaching many cells |
| Typical receptor | Often fast and ionotropic | Often slow and metabotropic |
| Typical timescale | Milliseconds | Hundreds of milliseconds to minutes or longer |
| Principal strength | Specificity—particular cells at a particular moment | Reach—many cells over a sustained interval |
| Closest cousin in the overview | Highly constrained paracrine signaling | Broader paracrine signaling; locally endocrine-like |
| Common examples | Fast glutamatergic and GABAergic signaling | Monoamines, acetylcholine, and neuropeptides |
These are tendencies rather than definitions. That qualification leads to the central conceptual question of the chapter.
19.3 Neurotransmitter or neuromodulator? A distinction of role, not molecule
It is tempting to read the table as a division between two kinds of chemical: the neurotransmitters, such as glutamate and GABA, in the wired column, and the neuromodulators, such as dopamine and serotonin, in the volume-transmission column. That division is convenient, and it reflects how the terms are often used, but it draws the boundary too sharply.
This reprises a principle that has appeared throughout the unit. In the overview, cortisol produced different effects depending on the receptor it encountered. In the previous chapter, glutamate produced rapid excitation through AMPA receptors, coincidence-dependent signaling through NMDA receptors, and slower biochemical effects through metabotropic glutamate receptors. The same principle applies here: “neurotransmitter” and “neuromodulator” are best understood as descriptions of function, not rigid categories of molecule. The same molecule can participate in either kind of signaling, depending on its receptor, its site and mode of release, and the machinery present in the target cell.
In the contrast developed here, fast neurotransmission carries a relatively precise point-to-point signal. Transmitter is released at a synapse, often acts through an ionotropic receptor, and produces a brief change in the conductance and voltage of the target cell. Neuromodulation changes how a neuron or circuit responds to other signals. It may alter excitability, neurotransmitter release, synaptic plasticity, or the relative influence of one input over another, often across broader regions and longer intervals. The distinction is not binary. Neuromodulation can occur at anatomically defined synapses, and transmitter released from a synapse can diffuse outward to extrasynaptic receptors. The terms identify different kinds of effect, not mutually exclusive molecular identities.
Two examples make the point especially clear.
Acetylcholine provides the cleaner case because its two principal receptor families have different names. At the nicotinic receptor—so called because nicotine binds to it—acetylcholine acts as a fast ionotropic transmitter. The nicotinic receptor is a ligand-gated cation channel in the same broad family as GABA-A and glycine receptors. When acetylcholine binds, the channel opens and depolarizes the cell within milliseconds. At the vertebrate neuromuscular junction, this mechanism converts neurotransmitter release from a motor neuron into rapid depolarization of the muscle fiber.
At the muscarinic receptor—named for muscarine, a mushroom-derived compound—the same acetylcholine acts through a metabotropic, G-protein-coupled receptor. Its effects develop more slowly and can persist much longer, altering ion channels, intracellular signaling, and neuronal excitability. Acetylcholine can therefore convey a rapid synaptic signal through one receptor family and alter the operating state of a cell through another.
Serotonin tells a similar story. It acts through more than a dozen receptor subtypes, nearly all of which are metabotropic G-protein-coupled receptors. Through these receptors, serotonin can produce slower changes in excitability, secretion, synaptic transmission, and circuit state. One serotonin receptor, however, is different. The 5-HT3 receptor is an ionotropic, cation-selective channel related structurally to the nicotinic acetylcholine receptor. When serotonin binds, it produces a rapid excitatory current. Its effect is functionally similar to the rapid depolarization produced by an AMPA receptor, even though the two receptors belong to different structural families. Serotonin, strongly associated with slow modulation, therefore also has a fast ionotropic mode.
The general lesson is that a signaling molecule’s biological effect cannot be inferred from the molecule alone. It depends on the receptor it encounters, the way it was delivered, and the signaling machinery of the target cell. When this chapter refers to “the neuromodulators,” it uses a convenient shorthand for molecules that predominantly participate in slower, state-setting forms of signaling, often through metabotropic receptors and often through relatively diffuse release. Those qualifications matter. The category is useful, but its boundary is porous.
19.4 The modulatory systems: small nuclei, brain-wide reach
Here is a fact that ought to be surprising and usually is not made surprising enough. The molecules that set the operating state of your entire brain — that make the difference between alert and drowsy, focused and diffuse, ready to learn and fixed — are manufactured by a handful of tiny clusters of neurons buried deep in the brainstem and basal forebrain. These are not large, distributed populations. They are small, discrete nuclei, some containing only tens of thousands of cells, and from these small sources arise axons that fan out to bathe vast territories of cortex and subcortex in their particular molecule. The architecture is the opposite of what the cortex taught us. A cortical pyramidal neuron addresses specific partners; a modulatory neuron addresses the neighborhood, and the neighborhood is most of the brain.
Take the clearest example first, because it sets the template for all the others. The locus coeruleus — the name means “blue spot,” for the pigment its cells carry — sits in the dorsal pons, and it is the brain’s only significant source of norepinephrine (also called noradrenaline). It is one of the smallest nuclei we will name: in the human brain it contains on the order of thirty to sixty thousand neurons, a rounding error against the brain’s eighty-six billion. And yet the axons of those few thousand cells project to nearly the entire brain — the whole cortex, the cerebellum, the hippocampus, the thalamus, the hypothalamus, the spinal cord. When the locus coeruleus fires, norepinephrine is released across enormous swaths of the nervous system at once, and the brain’s overall level of arousal and alertness shifts accordingly: low firing in sleep, moderate firing in calm waking, high firing in stress and vigilance. One of the brain’s tiniest structures sets one of its most global variables. That disproportion — minute source, brain-wide consequence — is the signature of a modulatory system, and it is what the rest of this section is variations upon.
The other major systems follow the same plan, each with its own molecule, its own nucleus or pair of nuclei, and its own characteristic influence.
Dopamine is manufactured chiefly in two adjacent midbrain nuclei: the substantia nigra (“black substance,” named like the locus coeruleus for a pigment its cells carry) and the ventral tegmental area beside it. These two send dopamine along distinct pathways with distinct jobs. The substantia nigra projects to the striatum of the basal ganglia and is central to the initiation and control of movement; its degeneration is the cause of Parkinson’s disease, whose tremor and rigidity follow directly from the loss of these dopamine cells. The ventral tegmental area projects to the striatum, the prefrontal cortex, and the limbic system, and is central to reward, motivation, and reinforcement — the pathway most directly implicated in addiction, and the one that will matter most in the next chapter. Dopamine is the modulator this book will lean on hardest when we come to learning, because dopamine is how the brain broadcasts the news that something better or worse than expected has happened. In 2009, Matsuda et al. reconstructed single nigrostriatal dopamine neurons whose axons formed enormous arbors within the striatum while sending almost no collaterals elsewhere. Their images reveal an important form of neuromodulatory architecture: a signal can be highly divergent within a selected target without constituting a brain-wide chemical bath. You can view the Journal of Neuroscience cover photo that shows the incredible reach of individual dopamine neurons.
Dopamine is often described as being released broadly into the forebrain, as though a small group of midbrain neurons poured the molecule over large regions of cortex and striatum. That image captures something important: dopamine neurons are few, their axons can branch extensively, and dopamine often acts beyond a single conventional synaptic cleft. But the metaphor becomes misleading if it suggests that dopamine is distributed uniformly or that every dopamine neuron broadcasts the same signal everywhere.
Specificity begins with the axons themselves. Individual dopamine neurons do not simply innervate the entire forebrain. Some preferentially target cortex, others particular portions of the striatum, and still others selected combinations of cortical and limbic structures. Matsuda and colleagues reconstructed nigrostriatal neurons whose axons formed enormous arbors within the striatum but sent almost no branches outside it. More recently, the POINTseq study mapped thousands of individual dopamine neurons and identified more than twenty-five recurring projection types. Many neurons contacted several regions, but the combinations were stereotyped rather than random. Dopamine’s long-range organization is therefore neither strictly point-to-point nor indiscriminately global.
There is further organization within cortex. Dopamine fibers vary greatly in density across cortical regions and layers, and optical imaging has revealed spatially distinct dopamine transients. Individual dopamine axons arriving in the same prefrontal territory can also respond differently to rewarding and aversive events. The phrase the mesocortical dopamine pathway therefore names a heterogeneous collection of axons rather than a single uniform signal.
Once dopamine has been released, it may still diffuse beyond a conventional synapse and activate extrasynaptic receptors. Yet this final step is local. Its reach is constrained by uptake, extracellular geometry, the positions of release sites, and the distribution and affinity of dopamine receptors. Even extrasynaptic receptors can be concentrated in specialized membrane microdomains.
The best description is therefore hierarchical: dopamine is broadly distributed at the level of the whole system, selectively routed at the level of projections, and locally diffuse at the site of release. It is not a uniform bath. It is a structured regional broadcast.
Serotonin comes from the raphe nuclei, a string of cell groups running up the midline of the brainstem through the medulla, pons, and midbrain. Like the locus coeruleus, the raphe project diffusely throughout the cortex and to most of the rest of the brain, and serotonin’s influence is correspondingly broad and hard to summarize in a word — it touches mood, anxiety, sleep, appetite, and aggression among much else. Its clinical importance is enormous: the selective serotonin reuptake inhibitors (SSRIs), the most widely prescribed class of antidepressants, work by raising serotonin levels at these diffuse synapses, and we will see exactly how in the pharmacology section.
Acetylcholine, which we just met as a molecule that is both transmitter and modulator, has its modulatory cell groups too. The principal one for the cortex is the basal nucleus of Meynert (and neighboring basal-forebrain nuclei), which supplies acetylcholine broadly to the cortex and is heavily involved in attention, learning, and memory — and is among the populations that degenerate in Alzheimer’s disease. A second cholinergic source in the brainstem (the pontine nuclei) projects upward and participates in regulating arousal and sleep–wake transitions. As a modulator, acetylcholine acts through the slow muscarinic receptors; the fast nicotinic story belongs to the wired mode.
The recurring pattern is clean enough to tabulate. Read down the table and the family resemblance is obvious: each is a small, deep source with diffuse reach, each acts predominantly through metabotropic receptors and volume release, and each is tied to a global function and to a disease that follows from its disruption.
| Modulator | Source nucleus | Major projections | Associated functions | Disorder of disruption |
|---|---|---|---|---|
| Norepinephrine | Locus coeruleus (dorsal pons) | Nearly the entire brain and spinal cord | Arousal, alertness, vigilance, stress response | Implicated in anxiety, depression, attention |
| Dopamine | Substantia nigra; ventral tegmental area (midbrain) | Striatum (movement); prefrontal cortex and limbic system (reward) | Movement, reward, motivation, reinforcement | Parkinson’s disease (nigral loss); addiction (VTA pathway) |
| Serotonin | Raphe nuclei (brainstem midline) | Diffuse throughout cortex and subcortex | Mood, anxiety, sleep, appetite, aggression | Depression and anxiety (target of SSRIs) |
| Acetylcholine | Basal nucleus of Meynert (basal forebrain); pontine nuclei | Cortex broadly (Meynert); arousal circuits (pontine) | Attention, learning, memory, arousal | Alzheimer’s disease (basal-forebrain loss) |
These pigmented nuclei connect back to something the lecture’s slides displayed: a structural MRI in which the substantia nigra and ventral tegmental area are visible as distinct dark patches in the living midbrain. The darkness is neuromelanin, a pigment built as a byproduct of dopamine and norepinephrine synthesis — the same pigment that names the nigra (black) and the coeruleus (blue). It is a small, satisfying fact that two of the brain’s most important modulatory nuclei advertise themselves by color, and that the color is a residue of the very molecules they manufacture.
Students meeting this material often trip over the fact that the locus coeruleus’s molecule has two names, and that a closely related molecule, adrenaline / epinephrine, has two more. The duplication is historical and geographic, not chemical. Adrenaline and noradrenaline are the older names, built from Latin (ad-renal, “next to the kidney,” where the adrenal glands sit atop the kidneys and secrete these molecules as hormones). Epinephrine and norepinephrine are the equivalents built from Greek (epi-nephros, “upon the kidney”), and they became the preferred terms in American usage, partly because Adrenalin had been registered as a trademark. The prefix nor- denotes the molecule lacking one methyl group relative to its partner: noradrenaline is adrenaline minus a methyl. For this book’s purposes, norepinephrine and noradrenaline are interchangeable names for the locus coeruleus’s modulator, and the kidney connection is a reminder of the overview’s point that the same molecules serve as hormones in the body and as signals in the brain — adrenaline is a circulating hormone of the fight-or-flight response and a neurotransmitter, depending only on where it is released and what reads it.
19.5 How a small nucleus helps orchestrate widespread brain activity
How can a relatively small population of cells influence such large portions of the brain? A useful analogy is an orchestra conductor. The conductor does not produce the music directly. Instead, by regulating tempo, dynamics, and coordination—and by directing particular sections at particular moments—the conductor changes how the musicians play together. Neuromodulatory nuclei exert a comparable influence. Their characteristic role is not to specify the detailed content of every perception, thought, or movement, but to help establish the conditions under which the circuits carrying that content operate.
These source nuclei have broad reach at the population level, but their projections are not indiscriminate. Molecularly and anatomically distinct populations of modulatory neurons send axons to particular regions or to recurring combinations of regions. Once an axon reaches one of its target territories, however, it may branch extensively and form numerous release sites. A small source population can therefore influence many cells. Broad influence is achieved through selective long-range routing followed by extensive branching within the selected targets.
Release within those territories can also occur in more than one way. Some transmitter is released at recognizable synaptic contacts; some spreads beyond a single synaptic cleft and reaches extrasynaptic receptors through volume transmission. This diffusion is local rather than brain-wide, and its reach depends on the arrangement of release sites, extracellular space, transporters, and receptors. At many targets, metabotropic receptors translate the chemical signal into changes in ion-channel activity, membrane excitability, synaptic strength, or gene expression. Because these effects depend on intracellular signaling cascades, they usually develop more slowly than ionotropic postsynaptic potentials and may outlast the release that initiated them. Selective projections, extensive local branching, extracellular diffusion, intracellular amplification, and persistence together allow a relatively small population of neurons to exert widespread—but still anatomically organized—effects.
The conductor analogy is useful because modulation changes how existing signals are processed. The conductor does not supply the notes; the musicians do. In the brain, fast excitatory and inhibitory communication is carried largely by ionotropic glutamate and GABA receptors. Neuromodulators can alter the gain of those responses, the balance of excitation and inhibition, and the probability that active synapses will be strengthened or weakened. The same sensory input can therefore produce a different response under different states of arousal, vigilance, stress, or motivation. This is the sense in which modulatory systems regulate faster synaptic processing rather than competing with it.
There is a further refinement. As the overview emphasized, a chemical signal need not mean only “present” or “absent.” Receptor affinity allows concentration to carry information. Many neuromodulatory neurons show tonic background firing punctuated by phasic bursts or pauses, creating different concentration profiles in their target regions. Affinity contributes to how these profiles are read, but it is not the only determinant: receptor kinetics and location, transmitter uptake, and intracellular coupling also matter. A low, sustained concentration and a brief, larger transient may therefore recruit receptor populations to different degrees and produce different consequences. In the next chapter, dopamine will provide an important example. Tonic and phasic dopamine signals have been associated with partly distinct functions, while phasic changes often carry event-related information—including reward-prediction-error signals in some dopamine populations—rather than simply announcing reward.
Neuromodulation also extends beyond neurons. Neuromodulators can act on astrocytes and vascular cells, altering vascular tone and modifying the relationship between neural activity and local blood flow. This connects the present chapter to the earlier discussion of neurovascular coupling. The familiar BOLD signal measured with fMRI is a hemodynamic consequence of neural activity, not a direct measure of neural firing. Changes in arousal or neuromodulatory state can therefore affect both circuit processing and the vascular response that accompanies it. Even the brain’s most widespread signals remain constrained by the metabolic demands of the tissue they regulate.
 ]
]
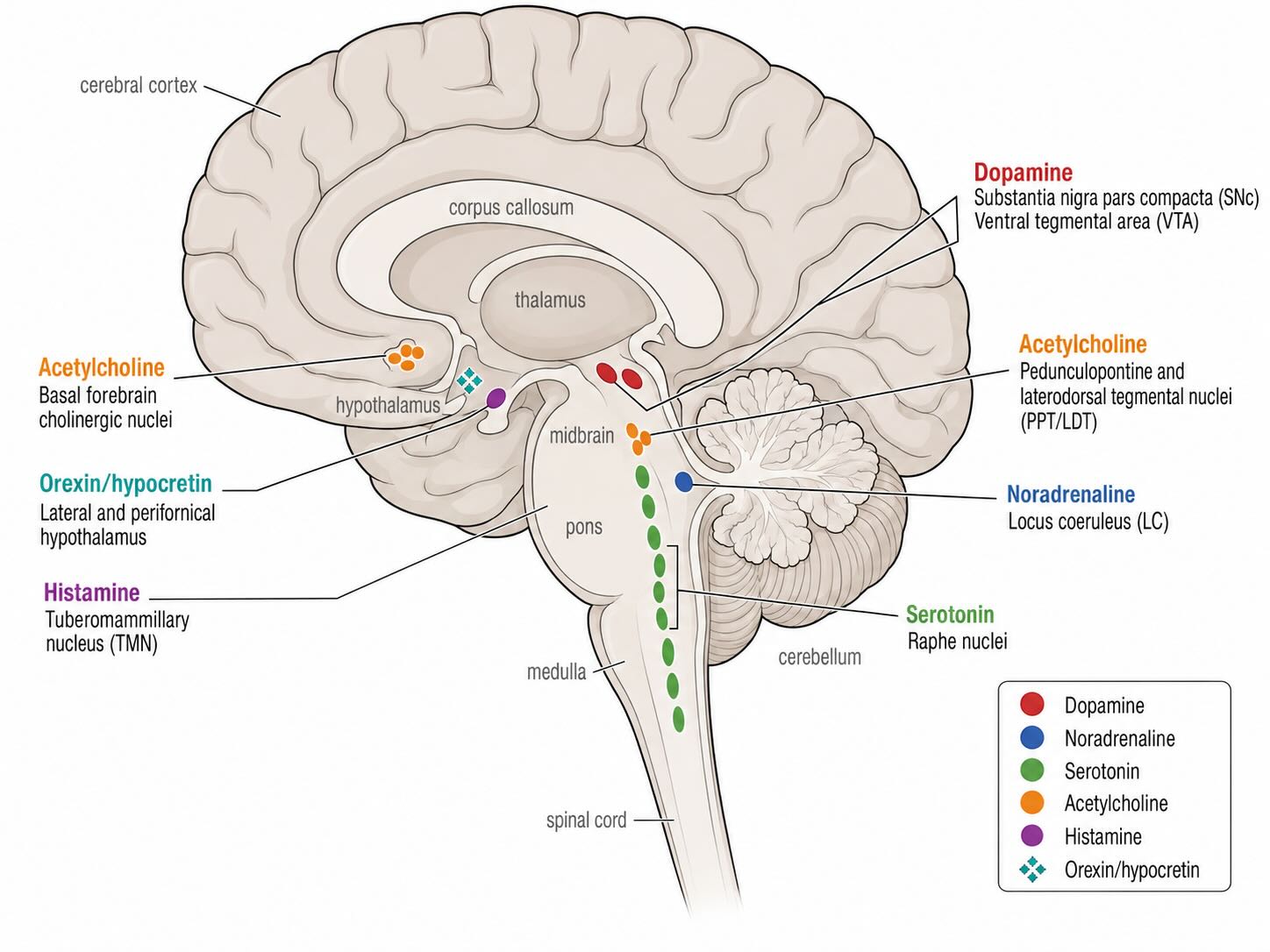
A striking feature of the brain’s neuromodulatory systems is how few neurons they contain. Each arises from a small, spatially restricted cluster of cell bodies, yet sends widely branching axons that influence large swaths of the brain. All counts below are rough, order-of-magnitude estimates from stereological studies, and the true figures remain debated.
- Dopamine — roughly 400,000–600,000 neurons, the great majority in the midbrain: the substantia nigra pars compacta (which projects to the dorsal striatum) and the ventral tegmental area (which projects to cortical and limbic targets), with a smaller contingent in the hypothalamic arcuate nucleus. These neurons are implicated in movement, motivation, and learning from outcomes, and are the population lost in Parkinson’s disease.
- Serotonin — on the order of a few hundred thousand neurons (often cited as approaching half a million), clustered in the raphe nuclei along the brainstem midline. The dorsal raphe holds the largest group. Serotonergic signalling is associated with mood, sleep, appetite, and pain modulation, and is the main pharmacological target of many antidepressants.
- Acetylcholine — on the order of hundreds of thousands of projection neurons, split between two main sources: the basal forebrain (medial septum, diagonal bands, and nucleus basalis of Meynert), which innervates the cortex and hippocampus, and a brainstem group (pedunculopontine and laterodorsal tegmental nuclei) supplying the thalamus. This system is associated with attention, arousal, and memory, and degenerates prominently in Alzheimer’s disease.
- Histamine — approximately 64,000 neurons per side (so on the order of 100,000+ in total), confined to the tuberomammillary nucleus of the posterior hypothalamus. Their projections reach the cortex, hippocampus, and much of the brain, and are associated with wakefulness and arousal, appetite, and thermoregulation.
- Orexin / hypocretin — roughly 50,000–80,000 neurons, located in the lateral and posterior hypothalamus. They densely target the other neuromodulatory nuclei (locus coeruleus, raphe, tuberomammillary nucleus, VTA), and are associated with the stabilization of wakefulness, feeding, and reward. Their loss causes narcolepsy.
- Noradrenaline — the smallest of these systems, with roughly 30,000–60,000 neurons (per-side counts cluster around 15,000), concentrated in the locus coeruleus of the pons, with a few smaller adjacent cell groups. Despite this tiny population, its axons reach the entire forebrain, cerebellum, and spinal cord, and it is associated with arousal, attention, and the stress response.
A note on the numbers. These estimates carry genuine uncertainty and vary across studies depending on counting method, the boundaries drawn around each nucleus, and how “projection neuron” is defined relative to co-localised GABAergic and glutamatergic cells in the same regions. The locus coeruleus range is a good illustration: per-hemisphere counts are fairly consistent (~15,000), but whole-nucleus totals are reported anywhere from ~30,000 to ~60,000. Treat the figures as conveying relative scale rather than exact values.
19.6 The pharmacology of the modulatory synapse
Neuromodulation matters well beyond the classroom. A large proportion of the drugs used to alter mood, arousal, motivation, or cognition — including antidepressants, antipsychotics, stimulants, nicotine, caffeine, and many drugs of abuse — act on modulatory signaling. Important exceptions act directly on glutamate or GABA receptors, so the division is not absolute. Even so, monoaminergic and cholinergic systems are unusually prominent drug targets because altering a relatively small, widely projecting system can change the operating state of many neural circuits at once.
The pharmacology becomes easier to organize if we follow the life cycle of a signaling molecule. A vesicularly released modulator must be synthesized from a precursor; packaged into vesicles; released into a synapse or the surrounding extracellular space; and bound by a receptor on its target cell. It must then be cleared so that the signal can end, either through a dedicated reuptake transporter or through enzymatic degradation. Synthesis, packaging, release, reception, reuptake, and degradation provide six potential points of pharmacological intervention.
Synthesis. Supplying more of a precursor can increase transmitter production. This is the logic of L-DOPA, a mainstay of treatment for Parkinson’s disease. Parkinson’s involves the progressive loss of dopamine-producing neurons in the substantia nigra. Dopamine itself crosses the blood–brain barrier poorly, but its immediate precursor, L-DOPA, crosses and can be converted to dopamine by surviving cells. The treatment therefore supplements a failing synthesis pathway from outside the brain.
Packaging. Transmitters must be loaded into vesicles before they can be released in the usual way. The vesicular monoamine transporter, or VMAT, moves dopamine, norepinephrine, and serotonin from the cytoplasm into synaptic vesicles. Reserpine blocks this transporter. Monoamines that cannot enter vesicles remain exposed to intracellular degradation, gradually depleting the stores available for release.
Release. Some drugs increase the amount of transmitter leaving the terminal. Amphetamine acts at several points in the monoamine life cycle, but an important part of its action is to disrupt vesicular storage, increase transmitter in the cytoplasm, and promote reverse transport through dopamine and norepinephrine transporters. The result is a large increase in extracellular monoamine concentrations.
Reception. A drug that binds a receptor and activates it is an agonist; one that occupies the receptor while preventing its activation is an antagonist. Nicotine is an agonist at nicotinic acetylcholine receptors — receptors named for their sensitivity to nicotine. Many antipsychotic drugs reduce signaling through dopamine D2 receptors, either as antagonists or partial agonists. Caffeine, by contrast, is an antagonist at receptors for adenosine, a locally produced modulator that accumulates during prolonged waking and contributes to sleep pressure. Caffeine reduces this influence by blocking adenosine receptors, thereby removing part of the restraint on arousal systems.
Reuptake. Blocking the transporter that recovers a released transmitter allows the transmitter to remain outside the cell longer. SSRIs — selective serotonin reuptake inhibitors — block the serotonin transporter and acutely increase extracellular serotonin. Their clinical effects emerge more slowly, as neural systems adapt to that initial change. Cocaine blocks the transporters for dopamine, norepinephrine, and serotonin. Its rapid blockade of the dopamine transporter contributes importantly to its reinforcing effects.
Degradation. Enzymes also limit transmitter availability, although they do not all operate in the same place. Acetylcholinesterase breaks down acetylcholine in the synaptic cleft, whereas monoamine oxidase, or MAO, metabolizes monoamines primarily within cells. MAO inhibitors reduce the metabolism of dopamine, norepinephrine, and serotonin and thereby increase their availability. These drugs were among the earliest antidepressants, and selective inhibition of the MAO-B form of the enzyme is also used to preserve dopamine signaling in Parkinson’s disease.
Once this framework is in place, an unfamiliar psychoactive drug can be approached with two initial questions: which signaling system does it affect, and which step in that system does it alter? Real drugs often act at several targets and across several timescales, but the framework provides a useful first approximation.
| Drug | Modulator system | Primary target step | Principal effect |
|---|---|---|---|
| L-DOPA | Dopamine | Synthesis | Supplies a precursor that crosses the blood–brain barrier and can be converted to dopamine |
| Reserpine | Monoamines | Packaging | Blocks vesicular storage and depletes releasable transmitter |
| Amphetamine | Dopamine, norepinephrine, and others | Release | Disrupts vesicular storage and promotes reverse transport |
| Nicotine | Acetylcholine | Reception | Activates nicotinic acetylcholine receptors |
| Caffeine | Adenosine | Reception | Blocks adenosine receptors and reduces a sleep-promoting influence |
| SSRIs | Serotonin | Reuptake | Block the serotonin transporter and increase extracellular serotonin |
| Cocaine | Dopamine, norepinephrine, and serotonin | Reuptake | Blocks monoamine transporters, producing a rapid rise in extracellular transmitter |
| MAO inhibitors | Monoamines | Degradation | Reduce enzymatic metabolism and increase transmitter availability |
The examples cluster around monoamines, acetylcholine, and adenosine. This does not mean that glutamate and GABA are pharmacologically untouched: benzodiazepines, many anesthetics, and ketamine are important counterexamples. Rather, diffuse modulatory systems provide especially powerful leverage over distributed brain states. A drug can alter how many circuits respond to their inputs without changing the content of every fast synaptic message.
19.7 How many neuromodulatory systems are there?
The figure in this chapter has already included histamine and orexin alongside the more familiar norepinephrine, dopamine, serotonin, and acetylcholine systems. Yet many introductory textbooks emphasize only the familiar four, sometimes presenting them as though they were the complete roster of the brain’s neuromodulatory systems. Histamine and orexin are therefore not exceptions that need to be appended at the end. They expose a more general problem: what qualifies as a neuromodulatory system, and how many such systems does the brain contain?
Part of the difficulty is that several different things can be counted. We might count chemical families, in which case dopamine is one modulator and serotonin another. We might instead count anatomically distinct projection systems. On that basis, dopamine neurons of the substantia nigra and ventral tegmental area could be treated as separate systems, as could the cholinergic neurons of the basal forebrain and those of the pontine brainstem. A transmitter name does not correspond neatly to one nucleus, one projection pattern, or one function.
The criteria for neuromodulation are also overlapping rather than absolute. An anatomical criterion emphasizes relatively small source populations whose axons diverge across large portions of the brain. A mechanistic criterion emphasizes volume transmission, metabotropic receptors, and effects that develop more slowly and persist longer than a conventional postsynaptic potential. A functional criterion asks whether a signal changes cellular excitability, synaptic plasticity, response gain, or the operating state of a circuit rather than conveying a narrowly specified moment-to-moment message.
The major diffuse systems satisfy several of these criteria at once, but none of the criteria is either necessary or sufficient. Acetylcholine and serotonin can activate ionotropic as well as metabotropic receptors. Neuromodulators can be released at conventional synapses as well as into extracellular space. Conversely, glutamate and GABA can produce slow modulation through metabotropic receptors. Adenosine is an important neuromodulator but does not arise from a compact nucleus with a brain-wide axonal projection. Endocannabinoids are synthesized on demand in postsynaptic cells and travel backward across local synapses. Thus, a neuromodulatory molecule is not necessarily part of a named diffuse projection system.
Histamine nevertheless fits the narrow diffuse-system model particularly well. Its neurons are concentrated in the tuberomammillary nucleus of the hypothalamus and project widely through the forebrain. Histamine acts through metabotropic receptors and contributes to the maintenance of wakefulness and arousal. The drowsiness produced by many first-generation antihistamines reflects their blockade of histamine receptors in the brain. By the same anatomical, mechanistic, and functional criteria used for the locus coeruleus and raphe nuclei, the histaminergic projection system belongs on the list. Its frequent omission is largely a matter of historical emphasis and pedagogical economy.
Orexin, also called hypocretin, satisfies many of the same criteria. Orexin-producing neurons form a relatively small population in the lateral and perifornical hypothalamus, but their axons reach widely distributed targets. Orexin acts through G-protein-coupled receptors and helps stabilize wakefulness while coordinating arousal with metabolic state and motivated behavior. Orexin neurons innervate several of the better-known modulatory populations, including noradrenergic, serotonergic, histaminergic, dopaminergic, and cholinergic neurons. The loss of orexin signaling is central to narcolepsy type 1.
Orexin is often placed outside the standard list because it is a neuropeptide rather than a small-molecule transmitter. Peptides are synthesized as larger precursor proteins in the cell body, packaged into dense-core vesicles, and transported down the axon. They are commonly taught as a separate chemical category, even when they perform functions that are plainly modulatory. Orexin was also identified only in 1998, long after the monoamine systems had become fixtures of neuroscience textbooks. Chemistry and history therefore help explain its placement, but neither provides a biological reason to deny that it is a neuromodulator.
There is consequently no single correct number of neuromodulatory systems. Under a narrow definition restricted to major long-range projection systems, the systems illustrated in this chapter provide a defensible introductory set — although even that number changes depending on whether related nuclei are combined or separated. Under the broader functional definition, the list expands to include many neuropeptides, purines, lipids, gases, and locally released signals. The standard textbook roster is useful, but it is a teaching selection rather than an exhaustive census. Histamine and orexin make the boundary visible because both satisfy the central criteria while falling outside the list that students are most often asked to memorize.
19.8 Looking ahead
We set out to explain the brain’s slow chemistry, and we can now describe it more precisely. Neuromodulation is not a single chemical family or a single anatomical arrangement. It is a mode of chemical influence in which a signal alters the responsiveness, plasticity, or operating state of cells and circuits. The major long-range systems commonly combine small source populations, divergent projections, metabotropic receptors, and some degree of volume transmission, but these are recurring features rather than a strict definition.
Their sources include the locus coeruleus for norepinephrine, midbrain nuclei for dopamine, the raphe nuclei for serotonin, basal-forebrain and brainstem populations for acetylcholine, the tuberomammillary nucleus for histamine, and the lateral hypothalamus for orexin. Their influence on human behavior is reflected in the large number of drugs that act on transmitter synthesis, packaging, release, receptors, reuptake, or degradation. Local modulators such as adenosine extend the same logic beyond the classic diffuse projection systems.
Three themes from the unit return here. First, neurotransmitter and neuromodulator describe roles rather than mutually exclusive kinds of molecule. Acetylcholine, serotonin, glutamate, and GABA can participate in fast transmission or slower modulation depending on their receptors, release conditions, and cellular targets. Second, the relationship between concentration and receptor affinity helps explain how tonic and phasic release can recruit different receptors and produce different effects. Third, the energetic theme reappears in the coupling among neural activity, vascular supply, and metabolic demand: changing the operating state of a circuit also changes the resources that circuit requires.
One further theme points directly toward the final chapter of the unit. In many reinforcement-learning settings, phasic activity in midbrain dopamine neurons carries information about how an outcome compared with expectation. Better-than-expected outcomes are often accompanied by a burst of activity, whereas worse-than-expected outcomes can produce a pause. This is not the only function of dopamine, but it provides a particularly clear example of how a delayed modulatory signal can guide synaptic plasticity.
The preceding chapter established the other half of this mechanism. When presynaptic activity is paired with postsynaptic depolarization, including a back-propagating action potential, recently active synapses can retain a short-lived biochemical record of their participation. This eligibility trace identifies synapses at which change is possible, but it does not by itself specify whether that change should be stabilized or in which direction it should proceed. A later dopamine signal can act on the trace before it decays, biasing the persistence and direction of synaptic change.
The fast synaptic events therefore establish which connections participated; the later modulatory signal supplies information about the outcome. Together they provide a biological solution to the problem of temporal credit assignment: how a brain changes the synapses responsible for an action when its consequences become known only afterward. Chapter 20 brings these two timescales together and examines how a synapse can use them to learn.