A plant does not have a brain, and this is not an oversight. A plant makes its living by standing still and turning sunlight, water, and air into sugar. Its food arrives where it grows. For an oak tree, the universe delivers — photons arrive, rain arrives, carbon dioxide diffuses to its leaves — and the adaptive response to most of these resources is to grow, slowly, in place. This should not be taken to mean that plants are inert. They track the sun, send roots toward water, and turn leaves toward light. But the timescale and mechanism matter: these are growth movements, played out over hours and days, achieved by differential cell expansion rather than by rapid behavior. A sunflower following the sun is not choosing a new action from moment to moment.
An animal makes its living differently. Animals are heterotrophs: they cannot build themselves out of sunlight, so they must find carbon that another organism has already fixed — eat the plant, or eat the thing that ate the plant. Once survival depends on going and finding, the animal faces a problem that the oak tree does not. It must move toward some things and away from others, and often do so now, on the timescale of seconds, because the food may be moving too, as may the thing that wants to eat it.
This is the deep reason animals have nervous systems and plants do not. A nervous system is what evolution builds when motility becomes the strategy. To move adaptively — toward food, away from harm — an organism must determine which way is which, and that requires sensing the world and converting what is sensed into movement. Sensing exists in the service of action. This is the thesis of the unit, and it will recur throughout: a sensory system is not a camera pointed at reality for its own sake; it is the front end of a control loop whose job is to produce the right movement.
There is an important sense, though, in which this unit has already started — in Chapter 6. In studying the hypothalamus, we watched the brain sense the state of its own body: detecting the osmolarity and sodium of the blood directly at the circumventricular organs, reading visceral and cardiovascular signals carried up the vagus nerve to the nucleus of the solitary tract, and measuring its own temperature with warm-sensitive neurons that are, quite literally, molecular thermometers. That was sensing too — but sensing turned inward, in the service of keeping the internal ocean within bounds. It has a name: interoception, the sensing of the body’s own internal state. It is the sensory front end of the homeostasis-and-allostasis story that has been the spine of this book.
That point deserves one further step, because it changes how what follows should be read. Interoception is not merely a regulatory loop operating beside behavior — it is very often what sets behavior in motion in the first place. Interoceptive signals are the prime movers. A falling blood-glucose level, rising plasma osmolarity, or cooling core can make an animal get up and do something: deploy its outward senses and muscles in search of a correction. Hunger sends an animal looking for food; thirst orients it toward water; cold drives it to seek shelter or sun. The hypothalamus does not just correct the internal variable through a hormone or a shiver. When the inexpensive automatic corrections are not enough, it recruits the entire exteroceptive and motor apparatus — the eyes, the ears, the legs — and directs it toward the world. This transition appeared in Unit II, even if it was not framed in these terms: the lateral hypothalamus energizing foraging, the switch out of consummatory rest into active search. The relationship between the two units is therefore not merely adjacent but causal. The inner senses generate the needs; the outer senses, the subject of this unit, are largely in the business of meeting them. Exteroception and movement are, to a large degree, the means by which interoceptive needs are satisfied.
This unit is therefore the other half of a pair — the half responsible for finding what the body needs. Having spent Unit II learning how the brain senses the body and generates its needs, we now turn outward to how it senses the world beyond the body — the surfaces, objects, sounds, and light beyond the skin — in order to act in it and meet those needs. That is exteroception, and it is what most people mean when they refer to “the senses.” The interoception/exteroception divide is not so much a new distinction as one already established: Unit II was, in large part, the interoceptive unit; this is the exteroceptive one. The seam will become sharper as the unit proceeds, while the somatosensation chapter will show where it also becomes biologically blurred.
When this book says that animals evolved senses “in order to” find food, the phrase is shorthand. Evolution does not plan or anticipate. No lineage looked ahead and decided that sensing would be useful. Rather, among organisms that varied, those whose movements were better tuned to their surroundings left more descendants. “In order to” compresses the longer statement that a trait was retained because of the fitness advantage it conferred. The short form will recur, but it should be understood in this longer evolutionary sense.
From single cells to a re-projected world
The link between sensing and moving is older than animals, neurons, and multicellularity. It is useful to see the link first in its simplest form, because the underlying logic does not change afterward; it is elaborated.
Phototaxis: a cell that swims toward the light
Even single cells sense and move. Consider a free-swimming alga such as Chlamydomonas. It has a light-sensitive patch — an “eyespot” — packed with rhodopsin-like photoreceptive proteins, and it has flagella that it beats to swim. The eyespot is positioned so that, as the cell rotates while swimming, it is alternately shaded and exposed by the cell’s own body. The photoreceptor signal is coupled almost directly to the beating of the flagella. The result is that the cell steers itself relative to light — phototaxis — turning toward an intensity favorable for photosynthesis and away from one that is too harsh.
The important point is how little machinery this requires, and how much of the essential organization is already present. There is a receptor, the eyespot, that transduces a physical variable, light, into a chemical and electrical signal. There is an effector, the flagellum. And there is a coupling between them such that the state of the world changes the movement. Receptor, coupling, effector. That is the logic of the nervous system in miniature, running in a single cell with no neuron anywhere in sight.
Chemotaxis: following an invisible gradient
Now consider an amoeba, or, for that matter, one of the body’s own neutrophils pursuing a bacterium. Here the relevant variable is not light but chemistry — the concentration of some molecule in the surrounding fluid. The cell reads the gradient across its own membrane, with more signaling molecules bound to receptors on one side than the other, and crawls up it toward food or toward the chemical signature of prey. This is chemotaxis, and again the architecture is receptor → internal signal → directed movement.
A fish detecting an amino acid leaking from a wound in the water is using a more elaborate version of the amoeba’s solution. The point of lining these examples up — alga, amoeba, fish — is not that they are the same, but that they sit on a continuum. The human sensory systems we will examine in this unit are not a discontinuous novelty. They elaborate a solution to a problem every motile cell has had to solve: which way should I go, given what I can detect?
The multicellular turn: why sensing at the surface is not enough
Multicellularity forces a genuinely new solution. For a single cell, the membrane is the sensor. The world reaches the cell directly, across its entire surface, and the cell responds. But a multicellular animal is a community of cells, most of which are buried in the interior, nowhere near the surface and certainly nowhere near a distant fox or an odor drifting on the breeze. The cells that have to act — muscle cells that move the body — are not the cells in contact with the world. The information therefore has to travel from the surface to the interior, and it has to arrive in a form the rest of the body can use.
The solution animals evolved is to dedicate specialized cells — sensory receptors — to the surface, and then to re-project the world inward: to build, inside the nervous system, an organized representation of what is outside, which the motor system can consult. The retina does not send “light” into the skull; it sends a patterned barrage of spikes that stands for the layout of the visual scene. The somatosensory system does not pipe “pressure” to the brain; it sends a signal that stands for where on the body surface contact occurred. The world is rebuilt, in neural code, on the inside.
The phrase re-projecting the world is useful because it identifies what to look for in every sensory system. In each one we will ask: what physical variable is being transduced, by what receptor, and how is the resulting map laid out inside the brain? Almost everything else is detail organized around that skeleton.
The shared plan of the vertebrate sensory systems
Everything above provides the deep background. We can now lay out the architecture that will recur, in one form or another, in the chapters on the body senses, vision, and hearing. This section is the canonical statement of that shared plan. The individual chapters will build on it rather than repeat it, so it is worth establishing clearly here.
A note on scope is needed before proceeding. This general plan is drawn from the “mechanical and visual” senses — touch, proprioception, hearing, and vision. The chemical senses, smell and taste, are being set aside temporarily. They are evolutionarily more ancient and wired differently — most conspicuously, smell is the one major sensory stream that does not route through the thalamus on its way to cortex in the way the others do. When we reach the chemical senses, that difference will be part of the explanation rather than treated as an awkward exception. For the senses that do share the plan, the following elements recur.
Receptors transduce one kind of physical variable
Every sensory stream begins with a receptor cell whose function is transduction: converting some physical feature of the world into the electrical currency of the nervous system. In many ways a sensory receptor cell is just a neuron where events in the world open the channels that cause its depolarization. What makes a photoreceptor a photoreceptor and a mechanoreceptor a mechanoreceptor is, quite literally, the nature of the environmental event that opens the molecular gate it carries in its membrane.
- Photoreceptors carry light-sensitive molecules (opsins) that change shape when they absorb a photon, triggering a cascade that changes the cell’s membrane potential.
- Mechanoreceptors carry ion channels that physically open when the membrane is stretched or deformed. Much of the molecular identity of these gates is now known: the Piezo2 channel, in particular, is the principal force sensor for both touch and proprioception in mammals [@coste2010piezo; @ranade2014piezo2touch; @woo2015piezo2prop]. The somatosensation chapter develops this finding; here the important point is that “a channel that opens when pushed” stopped being a metaphor and became a named protein within the past two decades.
- Thermoreceptors carry temperature-sensitive channels of the TRP family — TRPV1 for heat (the same channel capsaicin, the chili molecule, hijacks) and TRPM8 for cold (the one menthol hijacks) [@caterina1997trpv1; @mckemy2002trpm8].
The discovery of the molecular machinery for temperature and touch — TRP channels by David Julius, Piezo channels by Ardem Patapoutian — was recognized with the 2021 Nobel Prize in Physiology or Medicine. The significance of the award here is that it marks a real shift: the front end of sensation, long described only functionally as “a stretch-sensitive ending,” can now be described in terms of specific proteins. That is something a textbook written today can say that a textbook from twenty years ago could not.
Within a single modality there are usually submodalities, carried by different receptor types and kept partly separate all the way up. Vision divides into pathways more concerned with color and pathways more concerned with motion; touch divides into light touch, vibration, and the muscle sense. This branching recurs throughout the sensory systems.
A canonical route from receptor to cortex
Across the primary sensory systems, information is carried toward the brain through a sequence of neuronal populations. The terms first-order, second-order, and third-order neuron describe a neuron’s position in that sequence; they do not identify a particular kind of neuron. In the canonical arrangement shown in Figure 21.1, a first-order sensory neuron carries information from the periphery into the brain or spinal cord, where it contacts a second-order neuron. The second-order neuron then projects toward the forebrain, usually to a sensory-specific nucleus of the thalamus.
Neurons in the thalamus are conventionally called third-order neurons in this sequence. They project to the appropriate region of primary sensory cortex, terminating most densely in cortical layer 4. This organization is often described as a chain of relays, but that language can be misleading. At each stage, incoming activity is selected, combined, inhibited, amplified, and placed into a new relationship with other signals. The thalamus, in particular, does not merely copy sensory information from one location to another; it participates in transforming and regulating the activity that reaches the cortex.
Nor does the pathway end when activity reaches layer 4. Neurons in the superficial cortical layers, especially layers 2 and 3, communicate with other cortical areas. Neurons in layer 5 send output to subcortical structures involved in orienting, selecting, and producing action. Neurons in layer 6 project back to the thalamus. The canonical sensory pathway is therefore not simply an ascending line from receptor to cortex. It is embedded in a recurrent thalamocortical loop, with additional cortical and subcortical outputs that allow sensory activity to influence ongoing behavior. Other ascending routes, including pathways through the tectum, operate alongside this canonical thalamocortical route.
One final piece of anatomical bookkeeping is decussation: at some point in many sensory pathways, axons cross the body’s midline. Consequently, sensory information from one side of the body is usually represented most strongly in the opposite side of the brain. The location of the crossing differs among sensory systems—and sometimes among components of the same system—so it is more useful to ask where the second-order or later axons cross than merely to memorize that a pathway is “contralateral.”
The thalamus: many gateways, not one
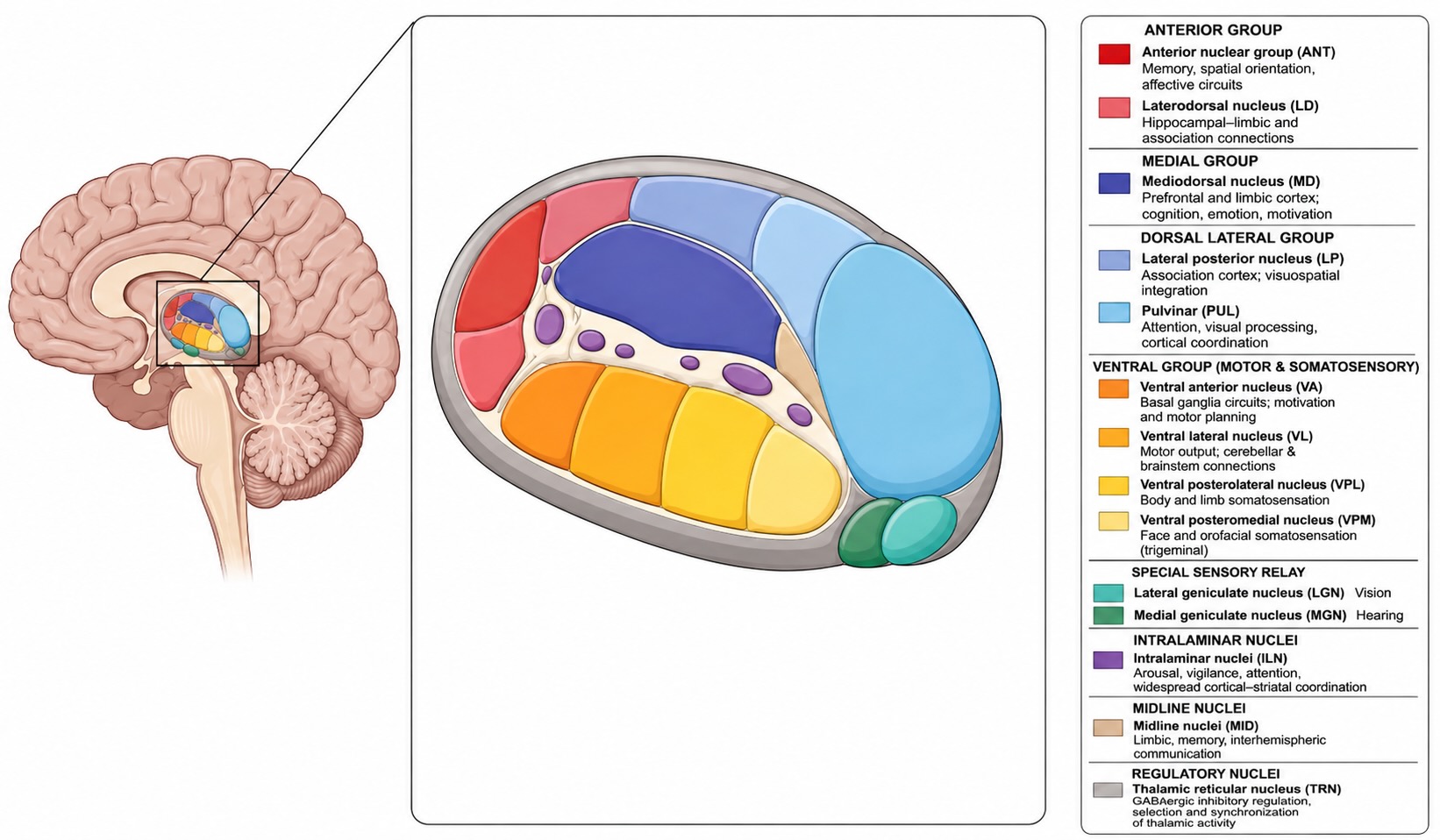
In Chapter 5, the thalamus was deliberately left as a closed box. We located it in the diencephalon, noted that it contains many nuclei, and moved on. That was enough for a first map of the brain. It is not enough in a unit on sensation, because the phrase “through the thalamus” hides both an anatomy and a circuit architecture.
The two thalami are paired, roughly ovoid masses of gray matter near the center of the forebrain. One lies on each side of the third ventricle, above the hypothalamus and medial to the internal capsule. The posterior thalamus expands into the pulvinar, with the lateral and medial geniculate bodies tucked against its posterior-inferior surface. Inside, a thin sheet of white matter called the internal medullary lamina helps divide the thalamus into anterior, medial, and lateral nuclear groups.
The point is not to memorize a complete thalamic atlas. It is to stop imagining the thalamus as one homogeneous relay. Thalamus names a structure, but the nuclei inside it belong to different loops.
Different nuclei, different loops
The thalamus can be classified along several overlapping axes. One asks which larger circuit a nucleus joins; another asks where its driver input originates—the input that supplies the principal information being relayed. This second classification uses the words first-order and higher-order in a specifically thalamic sense. Earlier, first-order, second-order, and third-order neuron described a neuron’s position in an ascending sensory sequence. Here, a first-order thalamic relay receives its defining driver input from below the cortex and is the first thalamic relay through which that stream reaches the cortex. A higher-order thalamic relay receives its defining driver input from cortical layer 5 and routes already-cortical information through the thalamus to another cortical area. Thus, neurons in the LGN can be third-order neurons in the retina-to-cortex sequence while the LGN is a first-order thalamic relay. The two labels answer different questions.
First-order thalamic relays: subcortical information entering the cortex. The best-known examples are sensory relays. They receive their driver input from a receptor organ or a subcortical sensory pathway and project toward the cortex, often to primary sensory cortex.
- The retina projects to the lateral geniculate nucleus (LGN), which projects to primary visual cortex, V1.
- Auditory pathways ascend through the brainstem and midbrain to the medial geniculate nucleus (MGN), whose ventral division projects strongly to primary auditory cortex, A1.
- Body and face pathways project to the ventral posterior thalamus: the VPL for the body and the VPM for the face, which project to primary somatosensory cortex, S1.
Geniculate means “knee-like”; the lateral and medial geniculate bodies are the principal visual and auditory relays. These are principal relays, not the whole sensory thalamus. Pain recruits additional lateral and medial thalamic territories, and taste has a thalamic relay that will be introduced in its own chapter. Smell is the conspicuous exception: olfactory information reaches primary olfactory cortex without first passing through an obligatory thalamic relay, although later olfactory processing does involve the mediodorsal thalamus.
Higher-order thalamic relays: cortical information routed through the thalamus. Higher-order relay circuits receive their defining driver input from layer 5 of one cortical area and project through the thalamus to another cortical area. Much of the pulvinar and parts of the mediodorsal thalamus participate in such communication among cortical regions. A signal can travel directly from one cortical area to another, but it can also travel by a parallel cortico-thalamo-cortical, or transthalamic, route. The layer-6 corticothalamic projection shown in Figure 21.1 is different: it provides feedback that regulates thalamic relay activity and reaches both first-order and higher-order relays.
Motor-related nuclei, especially the ventral anterior and ventral lateral nuclei, participate in loops carrying the results of basal-ganglia and cerebellar processing back toward frontal and motor cortex. They are not motor-command centers; they are nodes in recurrent systems for selecting, preparing, and refining action. Their designation as motor-related describes the larger circuits in which they participate, rather than a separate position in the first-order–higher-order classification. Subdivisions receiving their defining input from the basal ganglia or cerebellum can be first-order relays in the specifically thalamic sense.
The midline and intralaminar nuclei have broader connections with cortex and striatum and participate in brain state, behavioral relevance, and arousal. They should not be turned into a single arousal center. Waking state is produced by distributed interactions among thalamus, cortex, hypothalamus, basal forebrain, and brainstem.
These descriptions use several overlapping axes rather than dividing the thalamus into mutually exclusive compartments. First-order and higher-order identify the source of a relay’s defining driver input, whereas terms such as sensory, motor-related, and state-related identify the larger circuits in which it participates. Some nuclei contain subdivisions—or intermixed neuronal populations—that join different loops. That is not a problem with the classification. It is the biological point: the thalamus is a collection of circuit nodes, not one module with one function.
A relay is not a wire
Calling the thalamus a relay is not wrong. It becomes wrong only when relay is taken to mean a passive cable that copies an input unchanged. A relay identifies a route; it does not imply that nothing happens along the way.
Thalamic neurons can change the gain, timing, synchrony, and firing pattern of the signals they transmit. A nucleus can preserve the map carried by its input while altering when and how strongly particular parts of that map influence cortex. The LGN preserves the spatial organization of the retina, but it does not pass an untouched retinal photograph to V1.
A useful distinction is between driver and modulator inputs [@sherman1998drivers]. Drivers largely determine what a thalamic neuron represents; modulators alter how that information is transmitted. Retinal input is the principal driver of the first-order visual relay, while corticothalamic feedback, inhibition, and brain-state systems modify its transmission. In many higher-order nuclei, strong axons from cortical layer 5 can themselves serve as drivers. The result of one cortical operation can therefore become the input to another cortical area by way of the thalamus.
From primary sensory maps to actionable events
Primary sensory cortex: a map in layer 4
The principal first-order sensory nuclei project to distinct patches of cortex, and their axons terminate strongly in layer 4. A prominent layer 4 receiving dense thalamic input is therefore one anatomical signature of a primary sensory area:
- Vision → V1, in the occipital lobe.
- Hearing → A1, in the temporal lobe.
- Body senses → S1, in the postcentral gyrus.
This rule applies most cleanly to first-order pathways entering primary cortex. Higher-order thalamic projections can terminate in other cortical layers.
Primary sensory areas are organized as maps. S1 contains a somatotopic map of the body, V1 a retinotopic map of the visual field, and A1 a tonotopic map of sound frequency. In each case, relationships in a sensory feature space are preserved as spatial relationships on the cortical sheet.
Beyond primary cortex: objects, events, and possible actions
Primary sensory cortex preserves, in orderly maps, the dimensions that the receptors and thalamus deliver. The processing that follows does not simply relay these maps onward. It transforms them, step by step, from relatively local, receptor-derived variables toward relationships that are increasingly useful for behavior — and ultimately toward representations of objects and events in relation to the body and to possible action. That progression is the subject of this section. It is convenient to describe it as a hierarchy leading outward from primary cortex, and the description is a useful guide; but, as will become clear by the end, the actual circuitry is recurrent and the stages overlap more than any ladder suggests.
The classical vocabulary names the landmarks along the way. It divided cortex into primary sensory areas, unimodal (or modality-biased) association areas, and heteromodal or multisensory association cortex. The terms are historic but remain useful as a rough map. Association cortex is the broad label for cortex outside the primary sensory and motor areas: some of it stays biased toward one sense, some receives converging input from several, and much of it also carries information about posture, movement, memory, goals, and bodily state. Association cortex is therefore broader than multisensory cortex, and neither names one particular place. What follows is better understood as a change in what is represented than as a set of boxes.
The first steps beyond primary cortex remain modality-biased. Consider vision. Beyond striate cortex, extrastriate areas combine local orientation and contrast signals into representations of motion, depth, surface, and object form. A local motion signal is ambiguous — a short contour drifting across a small patch of retina is consistent with many global motions — so populations in these areas pool across space and orientation to recover the coherent movement of a whole surface, a variable that no single receptor reports. Higher auditory areas do the analogous work, combining frequencies and timing relationships into sound sources, voices, and other complex events. Secondary somatosensory and parietal areas combine signals across patches of skin and from proprioceptors to represent object shape and the changing configuration of the body. These areas are still meaningfully described as visual, auditory, or somatosensory. The progression here is not from sensation to thought; it is from local, receptor-derived variables toward relationships that behavior can act on.
At other points, information from the senses converges. The problem is not simply to add what the eyes, ears, and skin report. The brain must determine whether those signals were probably produced by the same event. A moving mouth and a voice should be combined when their timing and location agree; a voice coming from another part of the room should remain a separate event. When two cues probably share a cause, the more reliable cue can sharpen the estimate supplied by the less reliable one. When they probably have different causes, combining them would create an error. Multisensory integration therefore includes both binding signals that belong together and segregating signals that do not [@kording2007causal].
The posterior parietal cortex makes the connection to action especially clear. Neurons in different parietal territories combine visual, tactile, proprioceptive, vestibular, and eye- or head-position signals. They do not construct one neutral map of space. Different populations express locations in partly different reference frames and help transform a target from retinal coordinates into coordinates useful for looking, reaching, grasping, navigating, or defending the body [@avillac2005referenceframes]. A cup at one retinal location calls for a different reach when the eyes, head, or hand move. An approaching object matters differently when it is near the face than when it is far from the body. These are not sensory descriptions detached from movement; they represent the world in relation to possible action.
Parts of superior temporal cortex provide a complementary example. They respond to both the sight and sound of meaningful objects and moving agents, and they contribute to combining a speaker’s visible mouth movements with the voice arriving at the ears [@beauchamp2004multisensory]. The result is not merely a richer visual object beside a richer auditory object. It is a representation of one event — this animal moving, this person speaking, this object making that sound. Signals from frontal, limbic, and interoceptive networks can then alter the priority of that event according to what the animal is seeking, avoiding, or needing.
The familiar diagram — primary cortex → unimodal association cortex → multimodal association cortex — therefore captures a rough gradient but not the actual circuit, as the opening of this section anticipated. Cross-sensory influences can reach even nominally primary sensory cortex, while higher areas often retain strong modality biases [@bizley2007multisensory]. Integration also occurs outside cortex altogether — in the thalamus, in the superior colliculus, and along pathways that bypass primary sensory cortex entirely, as the later discussion of the tectum and alternative sensory routes will describe. There is no single cortical chamber in which completed sensory files are finally assembled into one internal picture. Instead, distributed and recurrent networks progressively construct objects and events in relation to the body and to action. The gradient is real as a summary of what each region emphasizes, even though the wiring that produces it is recurrent rather than serial: primary sensory maps preserve important dimensions of receptor input, while higher networks increasingly represent what is happening, where it is happening relative to the animal, and what that event affords.
Here is a claim that is genuinely clarifying, but that should also be held with some skepticism. Once the shared plan above is in place, the different sensory systems differ mostly in the modality their receptors transduce — and rather less in their downstream wiring than the separate labels might suggest. Cortex becomes “visual” or “somatosensory,” to a striking degree, because of the input it receives, not because visual or tactile specialization is fully built into the tissue in advance. We will see direct evidence for this in the somatosensation chapter, including an experiment in which cortex destined to be visual took on somatosensory organization after receiving somatosensory input during development. The implication is that receptors strongly specify modality, while cortical machinery is more general-purpose than its tidy functional labels suggest. The qualifier mostly matters. This is a strong claim doing real work, and its strongest form remains debated.
Sensory pathways are recurrent and parallel
Cortex talks back—and recruits inhibition
Neurons in layer 6 of cortex project back to thalamic relay nuclei, where they can alter the gain, timing, and state of the channels that originally drove cortex. Both thalamocortical axons ascending to cortex and corticothalamic axons descending from it give off branches to the thalamic reticular nucleus (TRN).
The TRN is a thin sheet of inhibitory neurons wrapped around much of the lateral surface of the thalamus. It is not the diffuse reticular formation of the brainstem, and it is not an ordinary relay nucleus. Its neurons do not project to cerebral cortex. Instead, the TRN receives branches from thalamocortical and corticothalamic pathways and sends GABAergic inhibition back into the thalamus. It is therefore positioned to regulate traffic between particular thalamic nuclei and their cortical partners.
Different sectors of the TRN are associated with different sensory and cortical systems. By inhibiting relay cells, the TRN can suppress some channels, sharpen contrasts among channels, coordinate thalamocortical timing, and participate in sleep rhythms. This makes it a plausible mechanism for selection: cortical systems can influence not only how they respond to an input, but how strongly that input is permitted to influence cortex in the first place.
In 1984, Francis Crick proposed that the thalamic reticular complex might control an internal attentional searchlight [@crick1984searchlight]. The proposal was speculative, but anatomically shrewd. The TRN is inhibitory, lies between thalamus and cortex, and is divided into sectors associated with different thalamic channels. Such a circuit could suppress some inputs while favoring others.
Later experiments supported important parts of the proposal. McAlonan, Cavanaugh, and Wurtz recorded from the LGN and adjacent visual TRN while macaque monkeys directed spatial attention into or away from a neuron’s receptive field. Attention enhanced responses in the LGN and produced an opposing change in the visual TRN, consistent with TRN-mediated regulation of the visual relay [@mcalonan2008gateway]. Wimmer and colleagues later trained mice to select between competing visual and auditory signals. Manipulating the visual TRN changed gain in visual thalamus and altered the animal’s ability to select the relevant modality, providing causal evidence that the circuit contributes to sensory selection [@wimmer2015selection].
The conclusion is stronger than “the TRN might be involved,” but weaker than “the TRN is the seat of attention.” Attention is a distributed control process involving frontal and parietal cortex, superior colliculus, pulvinar, basal ganglia, neuromodulatory systems, and sensory cortex. The TRN is one mechanism by which that larger system can impose selective inhibition on thalamic traffic.
Cortex can also communicate through the thalamus
Layer-6 feedback is not the only route from cortex into thalamus. Neurons in layer 5 send strong outputs to higher-order nuclei, which project onward to other cortical areas:
cortical area A → higher-order thalamus → cortical area B
This transthalamic route often runs beside a direct corticocortical projection. In a mouse brain-slice preparation, Theyel and colleagues interrupted the direct route from primary to secondary somatosensory cortex. Stimulation of primary cortex still drove secondary cortex until the higher-order somatosensory thalamus was also disconnected [@theyel2010corticothalamocortical]. More recently, Mo and colleagues inhibited layer-5 terminals from primary somatosensory cortex in higher-order thalamus while mice discriminated textures. Performance deteriorated even though the direct corticocortical route remained intact, and texture selectivity was especially disrupted in secondary somatosensory cortex [@mo2024transthalamic].
These findings do not make the direct cortical route unnecessary. They show that the transthalamic route can make an independent, behaviorally important contribution. Passing through higher-order thalamus may allow cortical communication to be adjusted by brain state, action-related signals, inhibition, and timing before it reaches the next cortical area. Exactly what transformation this route contributes remains unsettled.
The superior colliculus and pulvinar: a parallel visual road
A second kind of alternative route begins below cortex. The superior colliculus is not part of the thalamus; it is a layered midbrain structure involved in selecting targets and orienting the eyes and head. The retina projects directly to it, and the superior colliculus in turn projects to the inferior pulvinar, which projects to extrastriate visual cortex.
In macaque monkeys, Berman and Wurtz identified pulvinar neurons that received input from the superior colliculus and projected to motion-sensitive area MT, establishing a functional superior colliculus → pulvinar → MT route [@berman2010pulvinar]. This pathway does not replace the dominant retina → LGN → V1 route, but it provides extrastriate cortex with visual information that has not first passed through V1. It should not be inflated into a single explanation for blindsight, unconscious vision, or every rapid visual response. It is one well-established parallel path among several.
The pulvinar also participates in communication among cortical areas. In macaques performing a spatial-attention task, Saalmann and colleagues found that the pulvinar synchronized activity between interconnected visual cortical regions according to attentional allocation [@saalmann2012pulvinar]. The pulvinar can therefore participate both in an ascending route from the superior colliculus and in higher-order cortical communication. Different pulvinar territories participate in different loops.
A useful, deliberately simplified contrast is that the TRN can help regulate what passes through a thalamic relay, whereas the pulvinar can help regulate which cortical areas communicate effectively. Neither is an attention center. Together they make the larger point visible: the thalamus is not one gate to cortex but a collection of controlled crossings embedded in recurrent and parallel loops.
The canonical pathway remains useful: receptor → ascending pathway → first-order thalamic relay → primary sensory cortex. But it should be read as the spine of a recurrent network, not as a conveyor belt. Thalamic nuclei transform signals, cortex talks back, inhibitory circuits regulate passage, higher-order nuclei help cortical areas communicate, and parallel routes keep sensation tied to selection and action.
Tectum and cortex: reweighting, not replacement
Viewed from above, the brains of many fishes and amphibians present a striking image. The paired optic tecta form two of the most conspicuous structures in the brain, while the pallium of the forebrain appears comparatively small. The mammalian brain seems to reverse that relationship: the cerebral cortex dominates the surface, while the corresponding midbrain structure—the superior colliculus—is tucked beneath it. This comparison invites a simple evolutionary story. Perhaps an old tectal sensory system was gradually replaced by a newer cortical one, with some of its functions copied into primary sensory cortex.
That story is attractive, but it is not anatomically correct. The optic tectum and the superior colliculus are homologous structures in the midbrain (the mesencephalon vesicle of the neural tube shown in Figure 3.3) The cerebral cortex, by contrast, develops from the pallium of the telencephalon, at the anterior end of the forebrain. One structure did not transform into the other. Tectum and pallium were already distinct parts of the vertebrate brain long before the appearance of the six-layered mammalian neocortex.
There is a second caution. Lampreys, sharks, teleost fishes, amphibians, reptiles, and birds are living animals, not surviving ancestral stages. Each lineage has continued to evolve. A large tectum is therefore not simply a mark of a “primitive” brain. Its relative size reflects the sensory ecology and behavioral demands of the species. Nor does the absence of a mammalian neocortex imply the absence of a pallium. All vertebrates possess a pallium, although different lineages have organized and expanded it in very different ways.
Two sensory maps in one ancient vertebrate brain
Evidence from the lamprey makes the replacement story especially difficult to sustain. Lampreys belong to an early-diverging vertebrate lineage, but their brains contain both a well-developed optic tectum and a layered pallium. Recordings from the lamprey pallium have revealed a retinotopically organized visual region and an adjacent somatosensory region representing the head and trunk. Visual and somatic information reach these pallial territories through thalamic relays. Some retinotopically organized thalamic neurons also project to the optic tectum, preserving corresponding spatial organization across thalamus, pallium, and tectum. At the same time, the tectum retains its own circuitry for selecting orienting actions [@suryanarayana2020evolutionary].
The important result is not that the lamprey possesses miniature versions of mammalian V1 and S1. The exact homologies among pallial fields remain uncertain, and a recent cell atlas did not reveal the strong regional molecular arealization characteristic of mammalian cortex [@lamanna2023lamprey]. The important result is that mapped tectal and mapped pallial sensory systems already coexist in the same early-diverging vertebrate brain. Sensory cortex did not arise because an ancestral tectal map migrated into an empty forebrain. The vertebrate brain already contained two interacting systems for representing sensory events.
Shared computations do not imply shared anatomical origin
The more productive question is therefore not whether the tectum became cortex, but whether expanding pallial systems elaborated computations that were already present in the tectum. Here the answer is clearly yes—provided that “elaborated” is not mistaken for “directly descended from.”
Both systems organize sensory information into topographic maps. Nearby points in the visual world activate nearby locations in the tectum and in visual cortex. Both systems also perform feature filtering rather than merely relaying incoming signals. Tectal neurons can be selective for properties such as direction, speed, size, contrast, or looming motion. Both systems use inhibition to sharpen that selectivity and to make locations compete. In the lamprey tectum, for example, retinal excitation is followed by strong local and long-range inhibition that suppresses competing positions on the map. The result is a selection process: activity representing one possible target can win control of an orienting movement while rival locations are suppressed [@kardamakis2015tectal].
Related operations appear in sensory cortex: topographic organization, tuned receptive fields, surround suppression, recurrent competition, and transformations between sensory coordinates. These similarities are evolutionarily meaningful, but they do not establish that cortical circuits were copied directly from tectal circuits. Structural homology asks whether two regions descend from the same ancestral brain territory; the tectum and pallium do not. Circuit or cell-type homology asks whether particular neurons and connections share an ancestral origin; this remains unresolved for many comparisons. Computational analogy asks whether two systems solve similar problems; here the resemblance is unmistakable.
Primary sensory cortex is only part of the comparison
The superficial visual layers of the tectum have their clearest cortical parallel in early visual cortex. Both contain orderly maps of the visual field and extract behaviorally relevant structure from retinal input. Even here, however, primary visual cortex is not an enlarged superior colliculus. The principal mammalian route to V1 passes from retina to the lateral geniculate nucleus and then to cortex, in parallel with retinal projections to the superior colliculus. The two systems remain reciprocally connected and continue to influence one another.
The comparison becomes still less direct when the superior colliculus is considered as a whole. Its deeper layers combine sensory location with behavioral priority and potential movement. They help select where the eyes, head, body, or—in some species—the entire animal should turn. No single primary sensory cortex performs this full set of operations. In mammals, related functions are distributed across primary and higher sensory cortex, posterior parietal areas, frontal orienting systems, premotor cortex, basal ganglia, thalamus, and the superior colliculus itself. V1 may elaborate some computations found in superficial tectum, but the best cortical counterpart of the whole superior colliculus is a distributed sensory–attention–action network, not one primary area.
For the same reason, A1 and S1 should not be described as cortical copies of auditory and somatic maps in the tectum. Auditory and somatosensory information can reach the pallium through ancient ascending pathways of their own. The expanding cortex elaborated modality-specific analysis while the tectum retained a more compact role in locating consequential events and coupling them to action.
The name optic tectum can make the structure sound exclusively visual. Vision is usually dominant in its superficial layers, but the sensory content of the tectum varies with the animal’s ecological niche. In zebrafish, tectal neurons respond not only to visual and auditory stimulation but also to water movement detected by the mechanosensory lateral-line system [@thompson2016functional]. In weakly electric fish, visual and electrosensory signals occupy different tectal layers and converge in neurons whose dendrites cross those layers [@heiligenberg1987optic]. In pit vipers, an infrared-sensitive tectal layer lies beneath the visual layer, allowing heat emitted by prey to be registered alongside ordinary visual location [@hartline1978merging].
These examples suggest a broader definition of tectal function. The tectum does not contain a fixed list of “visual, auditory, and somatosensory” channels. It preferentially incorporates signals that can identify where a behaviorally important event is occurring and what orienting, approach, capture, avoidance, or defensive action that event affords. Water displacement, electrical fields, and infrared radiation can enter the system when those signals are useful to the species.
What did expanding pallial systems add?
Expansion of the pallium did not invent topographic mapping, feature selectivity, inhibition, or sensorimotor transformation. It greatly increased the number of separately differentiated sensory fields, the depth of recurrent and hierarchical processing, and the extent to which sensory responses could be shaped by learning, context, goals, and events that were not yet demanding an immediate movement. In mammals, modality-specific analysis became increasingly distributed across primary and higher cortical areas, while spatial priority and action selection became distributed across larger cortical–subcortical loops.
Even this distinction should not be turned into a simple contrast between a reflexive tectum and a thoughtful cortex. The tectum is influenced by forebrain, basal-ganglia, brainstem, and neuromodulatory systems, and its output can reflect behavioral state and context. Conversely, hierarchical pallial processing is not unique to mammals. A 2026 study of zebrafish found that the preglomerular complex sends visual and vibrational information to topographically organized pallial territories. Within the pallium, responses progress from relatively modality-specific activity to multimodal and nonlinear coincidence-detecting responses [@trinh2026hierarchical]. Yet developmental evidence indicates that much of the teleost preglomerular complex is derived from midbrain rather than from the diencephalic thalamus [@bloch2020nonthalamic]. A thalamocortical-like computational architecture can therefore arise from different anatomical components in different lineages.
The central evolutionary lesson is not replacement but reweighting. The vertebrate brain has long possessed both a tectal system and a pallial system. As pallial territories expanded, they elaborated several operations already conspicuous in the tectum and distributed others across larger recurrent networks. The superior colliculus nevertheless remained an active part of sensory processing and action selection. Evolution did not discard an old midbrain map when cortex appeared; it connected two ancient maps into increasingly elaborate loops.
The organizing axes of this unit, revisited
With the shared plan in place, the axis along which the unit is organized can now be stated explicitly — the axis introduced at the beginning and grounded in Unit II.
From interoception to exteroception
Recall the distinction: exteroception is sensing the external world — the surfaces, objects, sounds, and light beyond the body — whereas interoception is sensing the internal world — the state of the body’s own organs and tissues, including blood pressure, blood gases, gut distension, temperature, bladder fullness, and metabolic need. Interoception was developed in detail in the previous unit: the hypothalamus reading the blood at the circumventricular organs, visceral and baroreceptor traffic ascending through the vagus nerve to the nucleus of the solitary tract, and central thermometers in the preoptic area. Interoception is the sensory front end of homeostasis — how the brain learns that an internal variable has drifted so that corrective action can begin — and that was the central story of Unit II.
One striking point of contact between the two domains shows that the boundary is biological rather than bureaucratic: some of the same molecules do both jobs. The Piezo channels that mediate touch and proprioception, which we will examine in the somatosensation chapter, are also used to report blood pressure to the brainstem and to signal when the lungs and bladder are stretched. The same physics — a channel that opens when pushed — is read at the skin as the texture of the world, an exteroceptive signal, and in the wall of the aorta as the pressure of the body’s own blood, an interoceptive signal. The distinction is real, but it is drawn across a shared molecular toolkit.
The center of gravity in this unit is therefore the exteroceptive problem: how an animal builds a re-projected model of the world beyond its skin in order to act in it. The boundary is not perfectly clean, and the place where it blurs is biologically informative. The body senses — touch and limb position, but also temperature and visceral sensation carried through many of the same peripheral nerves — straddle the division. Sherrington’s classical account of the body senses included interoception as one of its three branches, alongside the outward-facing senses. The somatosensation chapter will use this overlap as an example of a point emphasized throughout the book: a distinction can be genuinely useful and still admit biologically meaningful exceptions.
From reactive to predictive sensing: the allostasis gradient
There is a second, subtler axis within exteroception, and it is especially important because it connects the senses to the homeostasis-to-allostasis story that runs through the book.
The exteroceptive senses differ in how far away the world they report is — and therefore in how much lead time they provide.
Touch is the sense of the world in contact with the body. Somatosensation reports events that are already happening to the body: a branch against an arm, the heat of a stove, the ground beneath a foot. There is essentially no lead time. A sense like this must feed reactive control — fast loops, reflexes, and corrections that begin before deliberation. This is homeostasis-style control in the sensory domain: detect the error, such as burning skin, and respond immediately by withdrawing. Much of the somatosensory system’s spinal-reflex machinery is built for this immediate-response regime.
Vision and hearing are senses of the world at a distance. Light and sound arrive from things that are not yet touching the body and often not yet affecting it directly — a predator across the clearing, a car approaching a crosswalk, a cliff edge ten paces ahead. A sense that reaches outward in space also reaches forward in time: it permits a situation to be detected before it becomes an emergency and therefore supports planning. This is the sensory substrate of allostasis — control by prediction, in which the brain forecasts a need and acts before the error arrives. In the language introduced in Unit I, the expensive brain is “buying prediction,” and the distance senses are part of what makes that purchase possible. Vision and hearing are, in this sense, time machines: they convert spatial distance into temporal warning.
The unit therefore picks up where the earlier interoceptive account left off. Unit II ended with the most inward and reactive sensing of all — the body monitoring itself. This unit begins just beyond that point, at the body surface: somatosensation, the sense of the world in contact with the body, still near the reactive end of the gradient. From there the unit moves outward to the distance senses that report what is there and what may happen soon. With that outward move comes a shift from reacting toward predicting, and from homeostatic correction toward allostatic preparation. Placed end to end, the two units form a single sweep from the inside out — from the chemistry of the blood, to contact at the skin, to light arriving from a distant hill. Across that sweep, the farther outward a sense reaches, the more future information it provides. The senses are not merely a list of input channels; they are arranged along an axis of temporal reach.
The unit ahead
This is one of the largest units in the book, deliberately so — sensation is where the brain’s relationship to the world is most concretely on display, and it will occupy a substantial share of the course. With the shared plan and the two organizing axes — internal versus external, reactive versus predictive — now in place, the individual sensory chapters can remain lean. Each will specify which physical variable its receptors transduce and how its particular maps and pathways are arranged, relying on this overview for the common scaffolding rather than rebuilding it.
We begin at the body surface with somatosensation, the touch-and-proprioception system. It is the natural opening for an exteroceptive unit that follows an interoceptive one: it sits at the boundary, remains near the reactive end of the gradient, and reveals where the interoception/exteroception distinction becomes biologically blurred. Many of the same peripheral nerves that carry touch also carry temperature and visceral signals. The chapter will also introduce cortical maps and the genuinely unsettled question of how fixed those maps are. From there we move outward into the distance senses, where sensation provides greater lead time and the brain increasingly shifts from reaction toward prediction.
With that framework in place, the unit begins with somatosensation, the sense of the body surface and the body in motion.
Reasonably settled:
- Animals have nervous systems because they are motile; sensing exists to guide movement. The receptor → coupling → effector logic is visible even in single cells (phototaxis, chemotaxis).
- The non-chemical vertebrate sensory systems share a plan: receptor → first- and second-order neurons → decussation → modality-specific thalamic nucleus → mapped primary sensory cortex. This pathway is embedded in recurrent thalamocortical and corticocortical networks. Beyond primary cortex, modality-biased and multisensory regions construct objects and events in relation to the body, the animal’s current needs, and possible action.
- The molecular identity of several key transducers is now known (Piezo for force, TRP family for temperature) — recognized by the 2021 Nobel Prize.
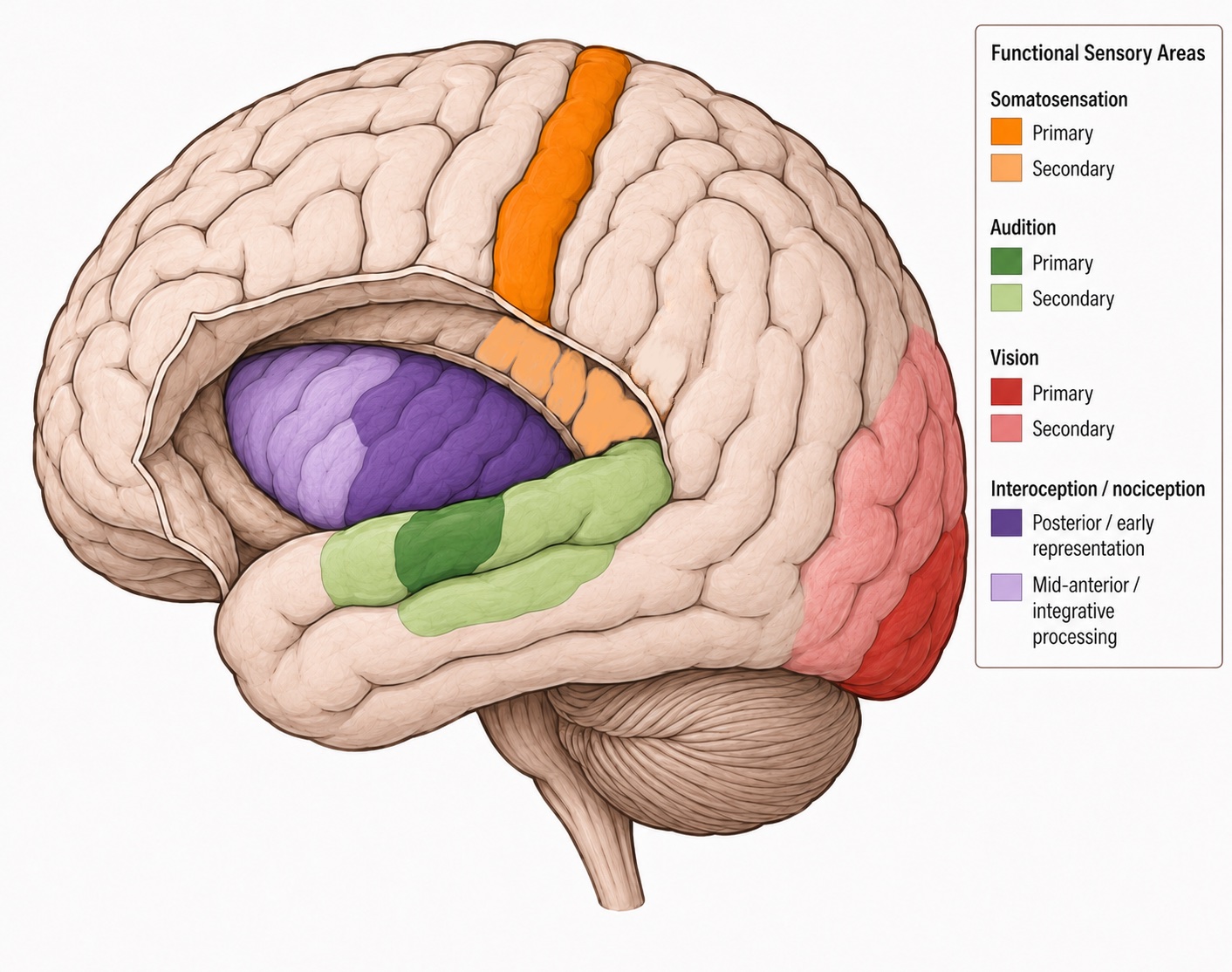
- The exteroception/interoception distinction is a real anatomical and functional seam, not merely a filing convenience: the two domains involve partly different receptors, pathways, and cortical destinations (with the insula prominent for interoception and the postcentral gyrus for exteroception). Interoception is the sensory front end of homeostasis — the central concern of Unit II — whereas this unit emphasizes exteroception; the body senses reveal where the boundary blurs.
Genuinely unsettled, and presented as such:
- What the massive cortico-thalamic feedback is for. Prediction, attention, gain control, sleep gating — all plausible, none established as the answer.
- Why sensory pathways decussate at all. Several hypotheses, none generally accepted (we take this up in the somatosensation chapter).
- How far “modality is just a receptor story” really goes — how general-purpose cortex truly is versus how much modality-specific structure is intrinsic. The developmental rewiring evidence is real but the strong form of the claim is still argued.
Despite these open questions, there is a great deal here that is well established. The shared sensory plan is one of the most robust organizing principles in neuroanatomy.