27 Vision III: Recognition, Awareness, and Action
What selective failures reveal about visual recognition, awareness, and action
Vision I showed that retinal output is already a transformed neural code rather than a copy of the image. Vision II followed that code through the optic chiasm, lateral geniculate nucleus, primary visual cortex, and the interacting networks beyond it. The visual-field map was preserved, but the code was progressively transformed: local contrast was recombined into orientation-selective responses, binocular differences supported depth, and different cortical populations placed different weights on motion, color, contour, texture, and surfaces.
This chapter approaches those transformations from the opposite direction. What disappears when a particular part of the system is damaged—and what remains? A person may lose color while retaining form, fail to recognize a familiar face while recognizing other objects, or guide a hand with visual information that cannot be reported as seen. These selective failures are informative because normal vision conceals its component operations. When the operations come apart, the construction becomes visible.
The method requires care. Strokes, tumors, traumatic injuries, and surgical resections do not respect the borders drawn in an atlas. A memorable patient may establish that a dissociation is possible without showing how commonly it occurs or identifying one unique mechanism. Conversely, the rarity of a clean syndrome does not make it an anecdote to be dismissed. When behavior, lesion anatomy, functional imaging, intracranial physiology, and causal perturbation converge, selective visual deficits provide some of the strongest evidence available for functional differentiation in the human brain.
27.1 A seamless visual field is constructed
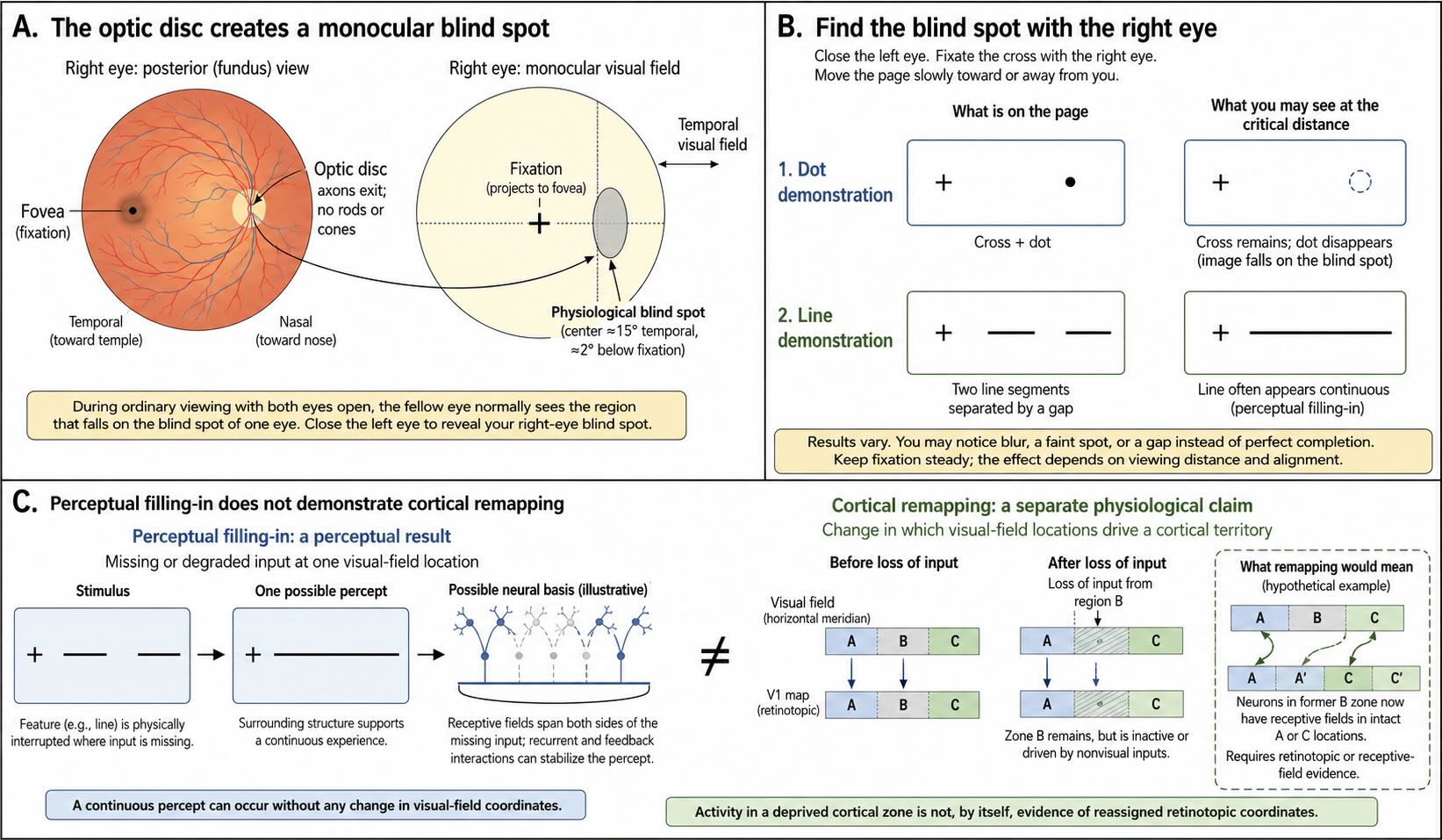
Each eye contains a region through which no image can be sampled. At the optic disc, ganglion-cell axons leave the retina to form the optic nerve, and no rods or cones occupy the exit site. The corresponding location in the visual field is the blind spot. It is a genuine absence of retinal input, yet ordinary visual experience contains no dark patch or empty hole.
Close the left eye and fixate a small cross with the right eye. Place a dot several centimeters to the right of the cross, then move the page slowly toward or away from the face without shifting fixation. At one distance the dot disappears as its image falls on the optic disc. Replace the dot with a gap between two aligned line segments and the line will often appear to continue through the missing region.
The disappearance of the dot can be explained by the absence of receptors. The apparent continuity of the line cannot. The visible ends of the line provide evidence that a contour continues through the unsampled region, and the visual system completes that contour. Textures and colors can also spread across the natural blind spot under appropriate conditions. This is perceptual filling-in: surrounding spatial structure contributes to what is experienced at a location from which no retinal signal arrives [@RamachandranGregory1991].
The term filling-in can sound as though a tiny painter inside the cortex colors a blank patch after the rest of the scene has been completed. That is not a necessary interpretation. Neural populations with receptive fields spanning the blind-spot representation can respond to contours and surfaces on both sides of it, and recurrent interactions can stabilize a representation that contains no experienced gap. The important result is phenomenological and computational: visual experience is assembled from relationships across space rather than read from a pixel-for-pixel retinal record.
Acquired visual-field defects are more complicated. Damage to the retina, optic nerve, optic radiations, or visual cortex can create a scotoma, whereas larger postchiasmatic lesions can remove a quadrant or hemifield. Some people report completion of lines, colors, or textures across an acquired blind region. Others experience blur, distortion, a sharp boundary, or partial awareness that something is missing. A hemianopic field does not generally behave as though it were simply an enlarged version of the natural blind spot [@Sergent1988]. The appearance depends on the cause, size, age, and location of the defect, the stimulus, and the remaining visual pathways.
The distinction matters because a seamless percept does not prove that missing retinal input has been restored. Monkeys with a small retinal lesion can show perceptual filling-in soon after the lesion without clear evidence that neighboring visual-field representations have invaded the deprived V1 territory [@MurakamiEtAl1997]. In humans with macular degeneration, activity found within the deprived portion of a visual map can often be explained by task demands, large receptive fields, feedback, or incomplete retinal loss rather than by large-scale rewiring of adult visual cortex [@BaselerEtAl2011].
Perceptual filling-in is the experience of continuity across a region with absent or degraded input. Cortical remapping is an anatomical and physiological claim that neurons deprived of their former input acquire a substantially different visual-field representation.
The first phenomenon is robust. The second can occur in limited forms, especially during development, but extensive adult remapping after a retinal lesion has not been demonstrated consistently. Changes in attention, feedback, receptive-field expression, and the unmasking of existing connections can alter activity without rebuilding the map. As in the body-map debate discussed in the somatosensation chapter, an altered percept should not be treated as automatic evidence of anatomical takeover.
27.2 How to reason from selective failures
The rest of the chapter moves from a natural gap in the retinal image to deficits produced by brain injury. Several different claims can be made from such evidence, and they should not be collapsed into one another.
The first claim is that a behavioral phenomenon exists. A person can lose color vision after cortical damage, perform above chance in a field described as blind, or fail to recognize familiar faces. A carefully studied individual may be sufficient to establish that a particular dissociation is possible.
The second claim concerns anatomy. Does the deficit recur after damage to a similar cortical territory, white-matter route, or functionally connected network? Lesion overlap can be highly informative, but vascular territories and neighboring functions can create correlations that are not themselves the mechanism.
The third claim concerns functional differentiation. Does a region respond more strongly or more selectively to one class of information than to appropriate controls? Functional MRI is particularly useful here because it samples much of the brain and can map the functional topography within an individual. Intracranial recordings sample far less tissue but measure local neural activity directly and with millisecond resolution.
The fourth claim is causal. Does damage, transcranial magnetic stimulation, or direct cortical stimulation alter the relevant percept or behavior? Causal evidence is powerful, but stimulation can affect axons and connected tissue as well as the neurons immediately beneath an electrode. No method arrives without qualifications.
The final claim is a broader theory of computation: that a region represents identity, computes motion, supports conscious awareness, or transforms visual coordinates for action. Such theories require the other levels of evidence, but they go beyond them. A deficit can localize a necessary contribution without revealing the complete algorithm.
The evidential strength will differ across the examples that follow. The relation between hMT+ and motion perception, ventral occipital cortex and color, ventral temporal cortex and faces, and several scene-selective regions is supported by unusually broad convergence. The exact pathway supporting a given instance of blindsight, the mechanism of Anton syndrome, and the cleanest version of the perception–action dissociation remain less settled. The aim is not to flatten all claims into equal skepticism. It is to place confidence where the evidence has earned it.
27.3 When one visual attribute disappears
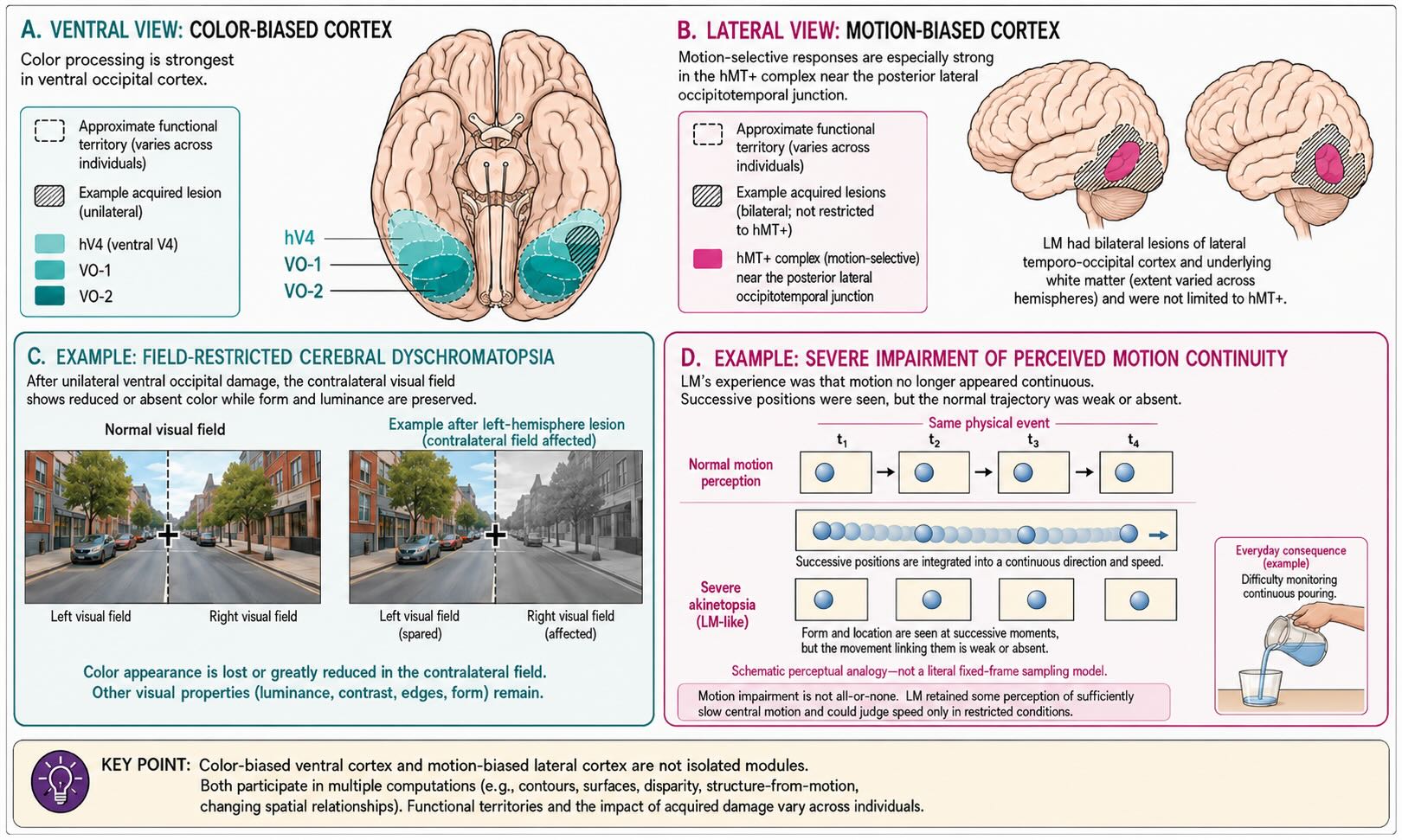
Vision II introduced hMT+ as a strongly motion-biased complex and ventral V4/VO-region circuitry as strongly biased toward color, form, texture, and surfaces. Selective losses of color and motion make those functional differences difficult to dismiss. They also show why the word selective is preferable to pure: a region can make a disproportionate contribution to one visual attribute while still participating in others.
27.3.1 Cerebral achromatopsia
Cerebral achromatopsia is an acquired disturbance of color perception caused by brain injury. It differs from inherited retinal color-vision deficiencies, in which one or more cone photopigments are absent or altered. In cerebral achromatopsia the retina may continue to encode wavelength differences, yet the person cannot experience, discriminate, or use color normally within the affected portion of the visual field.
Bilateral damage can leave the world largely gray or drained of normal color. Unilateral or smaller lesions can produce hemiachromatopsia or a quadrantic deficit, reflecting the retinotopic organization retained in ventral visual cortex. The loss can therefore be restricted to the contralateral visual field rather than following the boundaries of one eye. That pattern, together with preserved retinal function, strongly implicates postchiasmatic visual circuitry [@ZekiEtAl1991].
The relevant anatomy should not be reduced to one point labeled V4. A meta-analysis of 92 reported cases found the greatest lesion convergence in ventral occipital cortex, but lesion extent and impairment varied considerably, and many patients had additional spatial or form-related deficits [@BouvierEngel2006]. Human imaging likewise reveals several color-biased territories, including retinotopic hV4/VO regions and more anterior ventral temporal cortex. The secure conclusion is that normal color experience depends on a ventral occipital–temporal system containing reproducibly color-biased populations, not that one cortical area contains color and nothing else.
The convergence can be unusually direct. In one implanted patient, fMRI first localized an anterior color-biased site in the fusiform gyrus. Local field potentials recorded at that site were larger for chromatic than achromatic stimuli and differed among colors. Stimulation of the same electrode evoked a blue–purple percept resembling the color that produced the largest neural response [@MurpheyEtAl2008]. The result does not make that electrode the location of all color experience. It does connect, within one person, an imaging-defined functional bias, a directly measured neural response, and a stimulation-evoked percept.
27.3.2 Cerebral akinetopsia
Cerebral akinetopsia is an acquired impairment in perceiving visual motion. Its most famous description comes from patient LM, who had bilateral damage involving lateral occipitotemporal cortex. Motion no longer appeared continuous. Pouring liquid and judging approaching traffic became difficult because position seemed to change without the normally experienced trajectory between positions [@ZihlHeywood2015].
LM’s vivid experience should not be used as the definition of every motion deficit. Akinetopsia is rare, and impairment can depend on speed, direction, retinal location, contrast, and the complexity of the motion. Another carefully tested patient, for example, showed marked effects of target speed rather than a uniform absence of every motion signal [@HeutinkEtAl2019]. Some patients retain sensitivity to motion onset or can infer displacement from successive positions without experiencing normal movement.
The broader relation between hMT+ and motion has replicated far beyond one clinical case. Focal disruption of human V5/hMT+ with transcranial magnetic stimulation can transiently impair motion perception [@BeckersHomberg1992]. Functional imaging identifies hMT+ reliably across individuals despite anatomical variability, and motion-selective physiology, lesion effects, and stimulation all converge with the extensive macaque evidence reviewed in Vision II.
Color and motion therefore provide a useful pair, but not perfect mirror images. Ventral color-biased territories also analyze contours and surfaces. hMT+ contributes to disparity, structure from motion, and changing spatial relationships. Selective deficits reveal different weights within the visual system, not two sealed filters whose outputs are combined only after each has finished its private job.
27.4 What survives after V1 damage
Damage to primary visual cortex usually produces loss of vision in the corresponding portion of the contralateral visual field. A unilateral lesion can cause a homonymous scotoma or hemianopia; bilateral lesions can produce profound cortical blindness. Yet the destruction of V1 does not necessarily silence every visual influence on behavior.
The classic term blindsight refers to above-chance visual performance within a field that the person reports as blind [@WeiskrantzEtAl1974]. Depending on the patient and task, residual capacities can include detecting that a stimulus appeared, localizing it, discriminating motion direction or orientation, orienting the eyes, or adjusting a reach around an obstacle. These capacities are usually coarse and fragile compared with normal vision. They become visible because forced-choice testing asks for a response even when the person says that nothing was seen.
Blindsight is best treated as a family of findings rather than one preserved faculty. A person may succeed at motion discrimination and fail at form. Performance can depend sharply on contrast, speed, temporal frequency, and the exact borders of the lesion. Islands of spared V1 must be excluded where possible, but the absence of visible tissue on a structural scan does not guarantee that no geniculostriate neurons remain functional.
Several anatomical routes could carry visual information around the damaged portion of V1. Retinal signals reach the superior colliculus and pulvinar, as described in the unit overview and illustrated in Figure 21.4. The lateral geniculate nucleus also sends sparse projections directly to extrastriate cortex, and the intact hemisphere can contribute through interhemispheric connections. These routes coexist; the question is which one supports a particular residual behavior in a particular brain.
Diffusion imaging in a group of 17 adults with V1 lesions found that patients showing blindsight had a preserved pathway between the LGN and hMT+, whereas those without blindsight did not show the same preservation [@AjinaEtAl2015]. A later lesion-network study comparing patients with and without residual visual behavior found that connectivity with the ipsilesional medial pulvinar differentiated the groups [@KletenikEtAl2022]. The results need not describe one universal bypass. Motion discrimination, orienting, affective responses, and reaching can draw on different combinations of geniculate, collicular, pulvinar, extrastriate, and interhemispheric circuitry.
The word blindsight also raises a harder question: is the surviving processing entirely unconscious? In some experiments, patients perform above chance while denying any visual experience and while failing metacognitive tests that normal observers pass. Other patients report a vague feeling that something occurred—a shadow, flicker, pressure, or movement without ordinary seeing. When rating scales allow these weak experiences to be reported, performance can track degraded awareness rather than a complete absence of it [@OvergaardEtAl2008]. Other paradigms still find residual discrimination that differs substantially from normal conscious vision [@PersaudCowey2008].
The replicated core is therefore narrower and stronger than either extreme interpretation. Visual processing can survive major V1 damage and influence behavior without normal visual experience. The route and phenomenology vary. That variation is not a reason to discard blindsight; it is evidence that V1 is the dominant route into ordinary visual awareness without being the only route by which light can alter the brain.
The phrase can encourage a false anatomical picture: conscious vision through the LGN and V1, unconscious vision through the superior colliculus and pulvinar. The real architecture is more redundant. The LGN can reach extrastriate cortex without the destroyed portion of V1, the colliculus communicates with pulvinar and other motor-related structures, spared cortical islands may remain, and the intact hemisphere may contribute.
The behavior being measured matters. A pathway sufficient to bias an eye movement may not be sufficient to identify an object. Residual visual capacities should therefore be described task by task rather than assigned wholesale to one bypass channel.
27.5 When blindness is not acknowledged
Anton syndrome, or visual anosognosia, combines profound cortical visual loss with a failure to acknowledge that loss. A patient may insist that vision is intact despite colliding with objects, misidentifying what is in front of them, or offering confident descriptions unsupported by visual input. The descriptions may become confabulatory, although confabulation is not equally prominent in every case.
It is tempting to place Anton syndrome opposite blindsight. In blindsight, behavior shows more visual processing than the person reports; in Anton syndrome, the person reports more vision than behavior demonstrates. The contrast is memorable, but the syndromes pose different problems. Blindsight asks how visual information can still guide behavior after V1 damage. Anton syndrome asks why a person with severe visual loss fails to update beliefs about their own capacity.
Bilateral occipital injury can explain the blindness, but it is not sufficient to explain the anosognosia. Many people with cortical blindness know that they cannot see. Contemporary lesion and disconnection evidence points beyond visual cortex to disrupted interactions with parietal and frontoparietal systems involved in attention, monitoring, and the integration of current evidence with beliefs about the self [@MonaiEtAl2024]. Damage to visual association cortex and its long-range connections may prevent error signals from revising the person’s model of what they can do.
Confabulation can then be understood as one possible consequence rather than the primary mechanism. A person acts and speaks from a model in which vision is presumed to be available; when behavior fails, an explanation is generated from context, memory, or expectation. This account remains provisional. Anton syndrome is rare, lesions are often extensive, and no one network description yet explains every case.
Nothing in the syndrome requires the same mechanism that completes a line across the blind spot. Perceptual filling-in repairs a local spatial discontinuity using neighboring visual structure. Visual anosognosia concerns awareness of an impairment and the evaluation of one’s own performance. Both reveal that experience is constructed, but at very different levels of the system.
27.6 When form does not become an object
A person can have adequate acuity, fields, and elementary sensitivity yet fail to identify what is being seen. Visual object agnosia is a failure of visual recognition that cannot be explained by elementary visual loss, aphasia, generalized intellectual decline, or a global loss of knowledge about the object. The definition already signals a difficulty: recognition sits at the boundary among perception, semantic memory, language, and action.
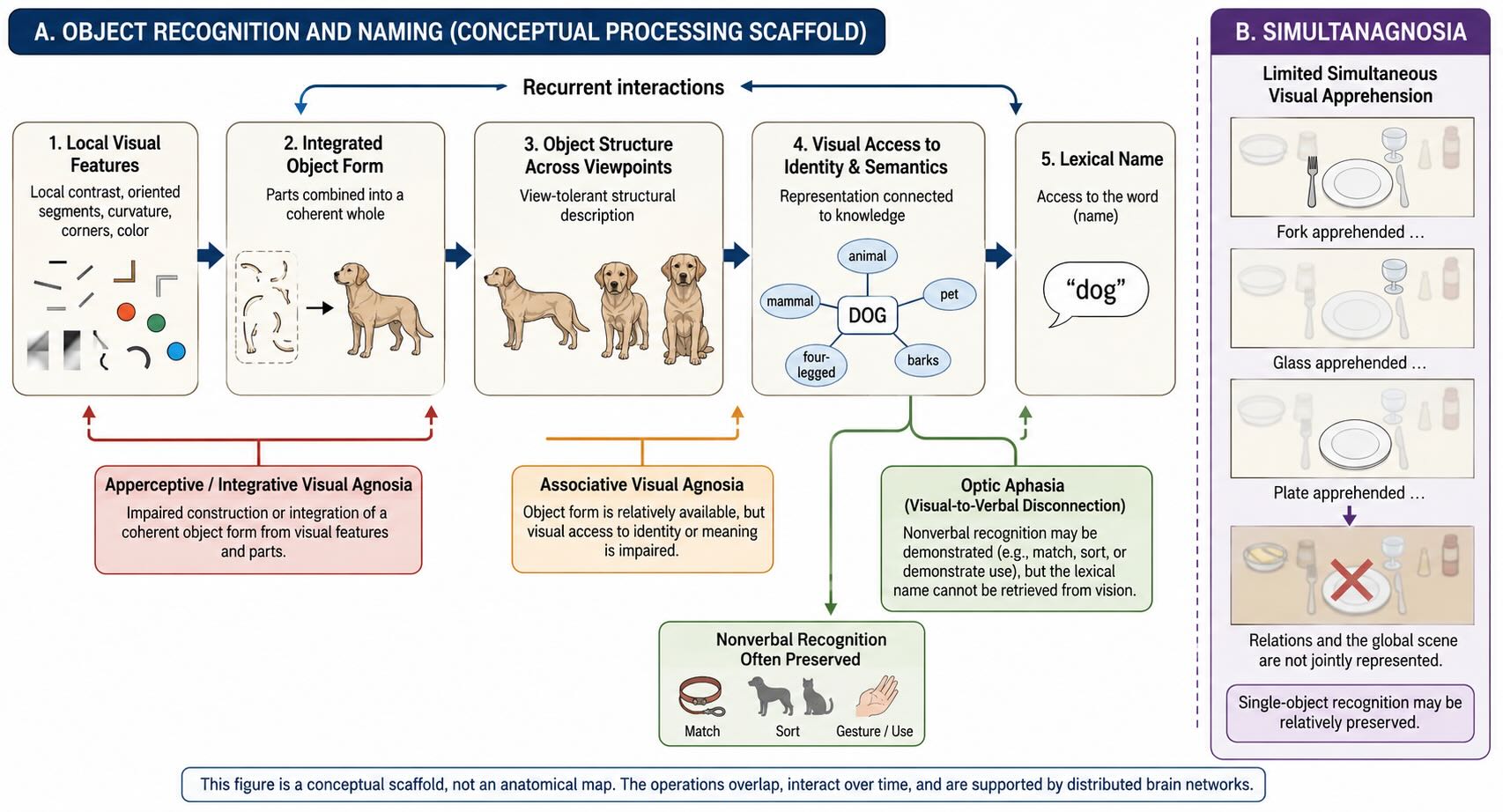
A useful conceptual sequence is:
local features → integrated shape → stable object description → identity and meaning → name
The sequence is not a claim that each step occupies one cortical station or that information moves only forward. It is a vocabulary for asking what a patient can still do. Can local contours be detected? Can the parts be combined into a coherent shape? Can two views be matched as the same object? Can the object be linked to what it is and what it is used for? Can its name be retrieved?
27.6.1 Apperceptive and integrative failures
In apperceptive agnosia, the failure occurs while a stable perceptual description is being constructed. A severely affected patient may struggle to copy a drawing, match shapes, or recognize an object presented from an unusual view. The eyes register local contrasts, but the visual system does not organize them into a sufficiently coherent object.
The category is itself heterogeneous. In integrative agnosia, local parts can be reported accurately while their relations are not combined into a global shape. Patient HJA, for example, could painstakingly identify details but was impaired when contours overlapped or when recognition required integrating several elements [@RiddochHumphreys1987]. Calling the problem “object recognition” is correct but incomplete; the immediate failure lies in organizing visual form.
27.6.2 Associative agnosia and access to meaning
In the classical associative form, copying and perceptual matching are comparatively preserved, yet a visually presented object cannot be identified or linked reliably to its meaning. Recognition may return when the same object is touched or heard. That cross-modal preservation shows that the concept itself has not disappeared; vision has failed to gain normal access to it [@McCarthyWarrington1986].
The contrast between apperceptive and associative agnosia remains useful, but real cases do not always occupy one clean box. A patient’s profile can change over recovery from a more perceptual toward a more associative pattern, and demanding perceptual tests can reveal deficits in apparently “pure” associative cases [@LarrabeeEtAl1985]. The distinction is best treated as a continuum of failure between the construction of form and access to identity, not as two indivisible diseases.
27.6.3 Neighboring syndromes are not interchangeable
Optic aphasia is primarily a visual-to-verbal disconnection. A patient may demonstrate an object’s use, sort it with related objects, or select a matching item while failing to name it from vision. That is not the same as lacking visual recognition altogether.
Simultanagnosia is an impairment in apprehending more than a very limited portion of a scene at once. A person may recognize one object but fail to represent the relationship among several objects or the global scene. It is usually associated with bilateral parietal or occipitoparietal injury and disturbed visual attention, often as part of Bálint syndrome. It belongs near the agnosias because the behavior can look like fragmented recognition, but the limiting operation is different.
Category-specific deficits require still more care. Some patients are substantially worse for living things than artifacts, or show the reverse pattern. Such cases helped motivate theories that semantic knowledge is organized partly by category, sensory attributes, and action associations [@WarringtonShallice1984]. Yet many famous living–nonliving dissociations are not purely visual. They can extend across words, sounds, and other modalities, indicating a semantic impairment rather than a visual agnosia. Animals also tend to share many visual features, while tools are strongly linked to characteristic actions. Perceptual similarity, familiarity, naming difficulty, and lesion location can all contribute.
The agnosias therefore do not reveal one final “object area.” They reveal several transformations by which contours become coherent forms, forms are matched across changes in viewpoint, and visual descriptions gain access to stored knowledge and names. Damage can interrupt different parts of that recurrent process.
27.7 Faces: reproducible territories and converging methods
Faces deserve separate treatment, not because every aspect of face perception is performed in one location, but because the evidential convergence is unusually strong. Selective impairments, nonhuman-primate physiology, fMRI, intracranial event-related potentials, and direct cortical stimulation all point to a highly structured system in ventral and lateral occipitotemporal cortex.
Prosopagnosia is a disproportionate impairment in recognizing faces. In severe acquired cases, a person may fail to recognize a spouse, colleague, famous individual, or their own reflection and instead rely on voice, hair, gait, clothing, or context. The patient may still know that the stimulus is a face and may retain some ability to judge gaze or expression. Other patients have broader visual-recognition deficits. Prosopagnosia is therefore a syndrome with several possible internal profiles rather than one perfectly uniform loss [@Farah2004].
27.7.1 From primate temporal cortex to the human ventral surface
The human work did not begin with a colored fMRI blob. Lesions producing prosopagnosia had long implicated posterior ventral occipitotemporal cortex, especially in the right hemisphere or bilaterally. In macaques, recordings by Gross and colleagues revealed inferotemporal neurons with striking preferences for complex forms, including faces [@GrossEtAl1972]. Perrett and colleagues then characterized temporal-lobe neurons responsive to faces and to particular views or features of faces [@PerrettEtAl1982]. These studies made a concrete prediction: human ventral visual cortex should contain localized populations that respond strongly and rapidly to faces.
The Yale–VA intracranial program provided direct human evidence. In 24 patients implanted with subdural electrodes for clinical seizure evaluation, faces evoked a large negative potential peaking at roughly 200 milliseconds—the face N200—at small regions of the fusiform and inferior temporal gyri. Scrambled faces, cars, scrambled cars, and butterflies did not evoke the same response at those sites. Electrical stimulation of the same regions frequently produced a temporary inability to name familiar faces [@AllisonEtAl1994Face].
Several points are important. The responsive sites were small and varied in exact anatomical position across patients. The N200 was a direct measure of local neural population activity rather than a scalp estimate or hemodynamic proxy. Its selectivity showed that face-sensitive processing had become spatially differentiated by about 200 milliseconds. The stimulation result added a causal perturbation: changing activity at those sites could transiently disrupt face recognition behavior. The electrophysiological response should not be equated with the complete representation of personal identity, but it identified tissue making a specialized contribution to the processing of faces.
27.7.2 Convergence within the same patients
The strongest feature of this research program was not merely that different methods pointed to roughly the same lobe in different samples. The methods could be compared within the same patients. In two implanted individuals, fMRI activation produced by faces was anatomically co-localized with the face-selective N200 recorded from subdural electrodes in right ventral extrastriate cortex; a similar left-hemisphere correspondence was present in one patient [@PuceEtAl1997]. Clinical stimulation mapping in these intracranial patients linked the same face-responsive ventral temporal territories to transient failures to identify or name familiar faces [@AllisonEtAl1994Face; @PuceEtAl1997].
That within-person convergence is unusually informative because the methods fail in different ways. fMRI provides broad spatial coverage but measures a vascular consequence of neural activity. Intracranial ERPs measure local neural activity directly and reveal its timing, but only from tissue that clinical needs happen to place beneath an electrode. Electrical stimulation can change perception or behavior, but the current spreads and can recruit fibers as well as local neurons. When a face-biased fMRI response, a face-selective intracranial potential, and a stimulation-induced failure of familiar-face identification converge in the same individual, no one method has to carry the argument alone.
Early fMRI studies from the same program then mapped face-sensitive extrastriate and fusiform responses in people without implanted electrodes [@PuceEtAl1995; @McCarthyEtAl1997]. The control conditions mattered. Faces and flowers each activated broad, partly overlapping bilateral fusiform territories when presented among nonobjects. When presented among familiar objects, however, faces isolated a smaller, predominantly right lateral fusiform response, whereas flowers produced no comparable focal response [@McCarthyEtAl1997]. The experiment therefore addressed selectivity rather than merely showing that visually complex stimuli activate ventral cortex.
Kanwisher, McDermott, and Chun independently replicated a focal fusiform response to faces, defined the region separately within individual participants, and tested it with independent contrasts [@KanwisherEtAl1997]. Their fusiform face area, or FFA, terminology provided a useful shorthand, and the individual functional-localizer strategy became standard. The historical sequence matters because the FFA was not a claim generated by fMRI in isolation. By the time the label became widely used, lesion evidence, primate physiology, intracranial ERPs, direct stimulation, and early fMRI had already converged on specialized face-responsive territories.
27.7.3 What has replicated
The basic fMRI result is reliable. Face-selective fusiform responses can be localized repeatedly within individuals, and the magnitude and spatial relationships of these responses show substantial measurement reliability [@McGuginGauthier2016]. Their exact coordinates vary, which is one reason that individual localizers outperform a single group coordinate. Variability in location is compatible with reproducibility in functional organization.
The early same-patient convergence has also been reproduced with newer recording methods. A later study compared fMRI, electrocorticographic responses, and electrical cortical stimulation within five patients and found 28 face-selective electrode locations identified by at least two modalities; no single method detected every site [@SanadaEtAl2021]. That result reinforces the original methodological lesson: agreement among measurements with different sensitivities is more informative than treating any one map as a complete inventory.
Independent stimulation studies have extended the causal evidence. In one well-known case, stimulation of face-responsive right fusiform cortex distorted the patient’s perception of a face while leaving nearby nonface objects comparatively unaffected [@ParviziEtAl2012]. Stimulation at other face-responsive ventral temporal sites has transiently impaired familiar-face naming or recognition [@AllisonEtAl1994Face]. Such effects differ across sites and stimulation parameters; they do not imply that one electrode contains a complete face. They show that activity in particular ventral temporal territories is necessary for normal face perception in that person at that moment.
Acquired prosopagnosia also need not follow only from direct destruction of the most face-selective fusiform patch. A lesion-network analysis assembled 44 reported lesions causing prosopagnosia. Twenty-nine intersected the right FFA directly, while the remaining lesions were located elsewhere; all were connected, in a normative functional-connectivity dataset, to the right FFA and to an additional frontal pattern [@CohenEtAl2019]. The result survived internal replication and predicted impairment in an independent lesion cohort. Lesion-network mapping is an inferential method based on connectivity measured in other people, and its details have been debated. Even so, the finding captures an important principle: a reproducible functional territory can be disrupted by destroying it, disconnecting it, or damaging another necessary part of its network.
The blood-oxygen-level-dependent signal is an indirect hemodynamic measure. That limits what fMRI alone can say about cellular mechanism and timing; it does not make functional localization empty. The usual claim is not that a patch of cortex is a sealed psychological “center.” It is that a reproducible territory shows a strong functional bias within a larger system.
Replication can therefore be asked at several levels. Can the territory be localized again in the same person? Does a similar topography appear across people and laboratories? Does selectivity survive new stimuli, tasks, and independent data? Does the response predict performance or impairment? Do lesions, intracranial recordings, or stimulation converge on the same territory?
Anatomical variability is not failed replication. A face-responsive patch that shifts substantially across individuals can still be highly reliable within each person and systematically located relative to nearby sulci and other functional territories. Individual functional localizers were developed precisely because averaging variable anatomy can blur a reproducible organization.
27.7.4 Not one face operation
Face perception is not one computation. The visual system must determine that a stimulus is a face, analyze its parts and configuration, distinguish one identity from another, recognize familiarity, and extract changeable information such as gaze, expression, and mouth movement. These demands recruit several functionally differentiated territories.
Posterior occipital face-responsive cortex contributes strongly to facial parts and early form. Fusiform territories place greater weight on coherent facial structure and information useful for identity. More anterior ventral temporal regions contribute to person-specific and semantic aspects of recognition. A lateral route through posterior superior temporal cortex is especially responsive to changing facial information, including gaze, expression, and movement. The borders are not absolute, and the regions communicate extensively [@HaxbyEtAl2000].
Recent lesion evidence strengthens the distinction between relatively static facial form and dynamic facial information. In 108 people with focal lesions, impairments in static and dynamic face perception showed a double dissociation associated with ventral and lateral temporal pathways [@PrabhakarEtAl2025]. The result supports causal differentiation beyond a single face system. Whether the lateral pathway should be formalized as a wholly separate “third visual pathway” is a broader theoretical question that remains under development.
The fine spatial organization is also richer than a map of isolated category boxes. Engell and McCarthy’s probabilistic work showed reproducible relationships and overlap among regions responsive to faces and biological motion [@EngellMcCarthy2013]. Their later intracranial study compared faces, isolated eyes, headless bodies, and flowers in 12 patients. Many electrodes responded more strongly to one or more of the socially relevant categories than to flowers, while only a minority responded exclusively to one of faces, eyes, or bodies. Crucially, adjacent electrodes often showed different response profiles, indicating spatially patterned sources rather than one undifferentiated “social” field [@EngellMcCarthy2014].
That result should not be summarized as either “the FFA is a sealed face module” or “ventral temporal cortex merely represents generic animacy.” The organization is distributed but highly structured. Faces, eyes, and bodies recruit overlapping tissue because their information is correlated in the world and because they belong to interacting perceptual tasks. Within that overlap, response maxima, timing, lateralization, and local spatial patterns differ. Coarse sampling can emphasize common activation; finer sampling reveals differentiation within it.
Faces may provide the clearest example of category-biased cortical organization because several developmental and ecological pressures point in the same direction. Faces are viewed repeatedly, usually near the center of gaze, share a constrained arrangement, and carry unusually consequential information about individual identity and social action. In that sense, faces have won a cortical lottery: they occupy especially reproducible functional real estate. That observation does not decide how much of the organization is genetically constrained and how much emerges through experience. It does make unconstrained learning and a uniformly distributed ventral stream implausible descriptions.
The most defensible conclusion is positive. Face perception depends on a reproducible set of cortical territories whose components place different weights on facial form, identity, movement, gaze, and expression. Those territories are neither isolated from the rest of vision nor interchangeable with it. The convergence of lesions, fMRI, intracranial ERPs, and direct stimulation is precisely what allows that statement to be made with confidence.
27.8 Scenes, landmarks, and finding one’s way
Faces are not the only category for which functional imaging has revealed reproducible high-level visual territories. Visual scenes recruit a set of regions including the parahippocampal place area (PPA) on the ventral surface, the occipital place area (OPA) near the transverse occipital sulcus, and retrosplenial/posterior medial cortex [@EpsteinKanwisher1998; @AguirreDEsposito1999; @DilksEtAl2013]. These regions are not copies of one scene representation. They place different weights on local layout, boundaries, landmarks, viewpoint, and the relation between the current view and a larger environment.
The fMRI topography has received causal support. Transcranial magnetic stimulation over the right OPA impairs scene discrimination more than face or object discrimination, whereas stimulation of nearby face- or object-selective regions produces different patterns of impairment [@DilksEtAl2013]. In an implanted patient, fMRI and intracranial EEG independently identified a place-selective region in the collateral sulcus and medial fusiform gyrus. Stimulation of that same region evoked vivid indoor and outdoor scene experiences, including familiar neighborhood views [@MegevandEtAl2014]. As in the color and face studies, the convergence within one person connects functional selectivity, directly recorded neural activity, and a stimulation-evoked percept.
A person who gets lost after brain injury does not necessarily have one unitary “place agnosia.” Topographic disorientation describes several failures [@AguirreDEsposito1999]. A patient with landmark agnosia may see a building clearly but fail to recognize it as the familiar landmark that anchors a route; ventral scene- and landmark-responsive cortex is especially relevant. In heading disorientation, familiar landmarks may be recognized, yet the patient cannot derive which direction to travel from one to the next; retrosplenial and posterior medial systems are often implicated. Parietal and OPA-region injury can disturb egocentric scene boundaries and the visual information needed to move through the immediate environment. Hippocampal and medial temporal damage can impair learning or retrieving the larger spatial relations among places.
The clinical taxonomy is imperfect because navigation normally combines all of these operations. Recognizing one’s house, knowing which way it faces, relating the current view to a map, and selecting the next turn are different accomplishments. A lesion can disrupt several at once. It is therefore inaccurate to treat every topographic deficit as the scene equivalent of prosopagnosia or to identify the PPA as a complete navigation system.
The broader lesson parallels the face work. Scene-selective fMRI territories are reproducible and behaviorally meaningful. Their functions become clearer—not weaker—when the system is divided into components and tested with lesions, stimulation, intracranial recordings, and tasks that separate local layout from landmark identity and large-scale navigation. The later unit on predictive maps will return to hippocampal and entorhinal contributions; here the emphasis is on the visual analysis that makes a navigable scene available to those systems.
27.9 Vision for action
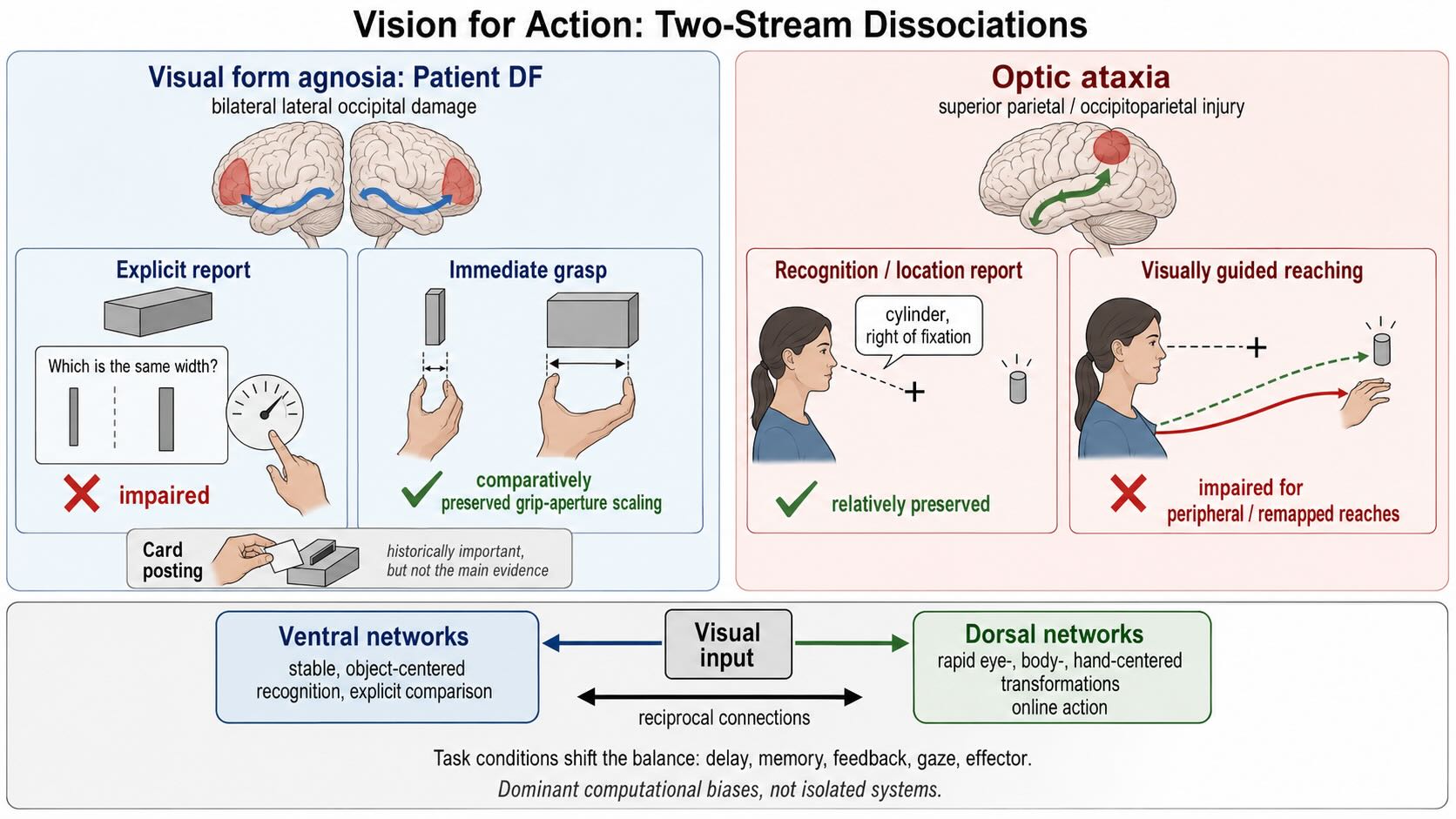
Vision II ended with interacting dorsal and ventral networks. The older shorthand called them “where” and “what” pathways. Goodale and Milner proposed a more consequential distinction: ventral networks contribute especially to enduring object-centered descriptions used for perception and recognition, whereas dorsal networks contribute especially to the rapid visual transformations needed to guide action [@GoodaleMilner1992; @MilnerGoodale2008]. Patient DF supplied the most influential evidence.
DF developed profound visual form agnosia after carbon-monoxide poisoning damaged bilateral lateral occipital and ventral visual regions. She performed poorly when asked to judge the orientation, width, or shape of visible objects. Yet some of her visually guided actions were strikingly better. Her grip aperture scaled to the width of an object as she reached to grasp it, even when she could not make an equivalently accurate perceptual report [@GoodaleEtAl1991].
The contrast suggested that the visual information required to identify or compare an object can dissociate from the information used to control a movement toward it. Online grasping requires the target to be represented relative to the eyes, hand, and current posture, with continual updating as the hand moves. Explicit recognition and comparison benefit from more stable descriptions that tolerate changes in viewpoint and can be retained or named. Those are different computational demands.
The famous card-posting task made the dissociation visually memorable. DF could not report a slot’s orientation accurately but could rotate a card while reaching and insert it through the slot. That result remains historically important, but later work has changed how much weight the task should carry. Healthy observers can perform posting by keeping the edges of the card clear of the slot rather than by explicitly matching the slot’s orientation. Posting errors follow obstacle-avoidance and biomechanical constraints that differ from those in perceptual orientation matching [@HesseEtAl2021]. The task therefore does not provide a pure comparison between two internal representations of orientation.
Grasp scaling provides a stronger example, but it too generated a productive dispute. Schenk reported that DF’s apparently normal grip scaling disappeared when trial-by-trial haptic feedback was removed and argued that tactile calibration might explain the classic dissociation [@Schenk2012]. Follow-up experiments designed to make haptic feedback uninformative still found that DF’s grip aperture varied with visually specified object size, although her performance was not normal in every respect [@WhitwellEtAl2014]. The challenge weakened the simplest version of the story without eliminating the broader difference between her perceptual judgments and online visuomotor control.
Complementary evidence comes from optic ataxia, usually after superior parietal or occipitoparietal injury. A patient may identify an object and describe its location yet misreach, misorient the hand, or shape the grasp poorly, especially for peripheral targets or when gaze and hand position must be transformed into a new coordinate frame [@PereninVighetto1988]. Optic ataxia is not the perfect mirror image of patient DF: deficits vary with visual field, delay, target visibility, effector, and lesion extent. It nevertheless demonstrates that normal recognition does not guarantee normal visual control of action.
The modern two-stream account should therefore be stated as a difference in dominant computational use, not a rigid separation between conscious perception and unconscious action. Ventral visual networks are especially important for relatively stable, object-centered descriptions that support recognition and explicit comparison. Dorsal networks are especially important for rapidly converting visual information into eye-, body-, and hand-centered parameters for online action. The streams exchange information continuously. Delays, memory demands, feedback, peripheral versus central presentation, and the kind of movement can change the balance between them.
That formulation preserves what the patient evidence established while discarding the cleanest textbook exaggerations. DF did not prove that the dorsal stream acts without ventral input in every circumstance. The posting task did not isolate orientation as neatly as once assumed. Yet the contrast between DF, optic ataxia, and ordinary behavior still shows that seeing an object for recognition and using visual information to act on it are not one indivisible operation.
27.10 Coda: the transformations can fail separately
The visual system does not begin with a picture and then add color, motion, identity, and meaning as optional labels. The retina transforms light before the optic nerve is formed. V1 and extrastriate cortex preserve spatial relationships while changing the code. Later systems extract partially separable information about surfaces, movement, objects, faces, scenes, and possible actions. Recurrent interactions normally bind these contributions into a visual world that feels immediate and unified.
The selective failures in this chapter reveal that unity from different directions. Filling-in shows that spatial continuity is constructed from context. Achromatopsia and akinetopsia show that color and motion depend on differentiated cortical circuitry. Blindsight shows that visual information can influence behavior without normal V1-mediated experience. Anton syndrome shows that losing vision and knowing that vision has been lost are not the same neural event. The agnosias separate the construction of form from access to identity, meaning, and names. Prosopagnosia and topographic disorientation expose reproducible high-level visual systems for faces and scenes. DF and optic ataxia reveal different demands of recognition and online action.
These phenomena should not be forced into one common mechanism. The cortex does not “fill a void” in the same way when it completes a line, confabulates during anosognosia, recognizes a face, or guides a grasp. The common principle is architectural: visual experience and visually guided behavior depend on multiple transformations that can fail separately.
That principle returns the unit to its starting point. Vision matters because it permits action before contact. A face is not only recognized; its gaze and expression alter what another person may do. A scene is not only seen; it provides boundaries, routes, shelter, and obstacles. Motion specifies trajectories. Color helps separate surfaces and identify useful objects. The next unit follows the control loop across the seam from sensing into doing: how targets are selected, movements are specified, errors are corrected, and the body acts on the world that vision has made available.
Reasonably well established
- Visual experience is not a pixel-for-pixel copy of retinal input. Contours, colors, and textures can be perceptually completed across the natural blind spot.
- Perceptual filling-in does not by itself demonstrate large-scale cortical remapping after retinal or cortical injury.
- Cerebral achromatopsia and akinetopsia are genuine acquired disorders. Ventral occipital color-biased territories and hMT+ make strong, differentiated contributions to color and motion perception, respectively.
- Residual visual processing can survive extensive V1 damage and can influence forced-choice judgments, orienting, and action without normal vision.
- Visual object agnosias can interrupt different stages between the integration of form and access to identity, semantic knowledge, and names.
- Face-responsive ventral temporal territories are reproducible. Lesions, fMRI, intracranial ERPs, and direct stimulation converge on their importance, including convergence of these methods within the same patients.
- Face perception is internally differentiated: ventral territories place strong weight on facial form and identity, while lateral temporal territories place greater weight on dynamic information such as gaze, expression, and movement.
- PPA, OPA, and retrosplenial/posterior medial cortex form reproducible but differentiated components of scene perception and navigation-related processing.
- Visual information used for explicit recognition can dissociate from information used for online action. Dorsal and ventral networks nevertheless interact extensively.
Established in outline but not captured by one simple mechanism
- Acquired scotomas vary in whether and how they support filling-in; they should not all be treated as enlarged natural blind spots.
- Blindsight is not supported by one universal bypass pathway. LGN–extrastriate, collicular–pulvinar, spared V1, and interhemispheric routes can contribute differently across patients and tasks.
- Residual vision after V1 damage ranges from performance without reported awareness to weak or degraded visual experience. No single phenomenological description fits every case.
- Anton syndrome requires more than occipital blindness, but the precise combination of visual, parietal, frontal, and disconnection effects producing visual anosognosia remains unsettled.
- The apperceptive–associative distinction remains useful, although many patients fall between the classical endpoints and category-specific deficits may be perceptual, semantic, or both.
- The functional organization of face-responsive cortex is strongly constrained and replicable without being one sealed module. The relative contributions of developmental experience, connectivity, and evolved bias remain active questions.
- The DF literature continues to support a perception–action distinction, but haptic calibration, task strategy, delay, visual field, and lesion extent limit the strongest claims. Card posting is no longer a clean test of visual orientation processing.
The productive conclusion is not that localization failed or that every operation is everywhere. The visual brain contains reproducible functional differentiations embedded within interacting networks. Selective deficits reveal those differentiations; variability across lesions, tasks, and pathways reveals how the networks are connected.