4 The Embodied Brain
The brain in a body
We ended the last chapter by noting that a patterned neural tube is a controllable tube. Rather than a uniform cylinder of tissue, the tube is carved by regulatory genes into specific territories with distinct addresses. These addresses mark the origins of neural structures that will eventually execute highly specialized functions. That is where we left the developing brain—built, regionalized, and ready.
Ready to do what, exactly? That is the central question of this chapter. Most textbooks follow early development with an immediate anatomical survey: pointing out the thalamus, the cerebellum, the four lobes, and asking you to memorize the gyri. We will conduct that structural survey in Chapter 5. Before we catalog the anatomy, however, we need to establish the fundamental operating principles that govern those structures. Before naming the parts, we must first understand the brain as a biological control system designed to drive allostasis and maintain internal balance.

Ascending Interoception (Blue pathways): The brain is exquisitely sensitive to the physical state of the internal organs. Afferent signals detailing conditions in the gut (originating in the enteric nervous system) and blood pressure (via cardiovascular baroreceptors) travel upward, primarily via cranial nerve IX and the vagus nerve (CN X). These ascending sensory tracks converge on the nucleus of the solitary tract (NTS), the brainstem’s principal hub for interoceptive data. Concurrently, the brain monitors peripheral immune responses neurally via the vagal inflammatory reflex.
Humoral Sensing & Secretion (Purple pathways): The central nervous system directly monitors non-neural signals in the blood. Specialized circumventricular organs (such as the area postrema) lack a typical blood-brain barrier, allowing them to act as direct blood sensors. They actively sample the systemic circulation for shifts in osmolarity, microbial metabolites, and immune cytokines. However, this communication is not one-way: the brain is itself a secretory organ, releasing its own neuroendocrine hormones into the systemic circulation to modulate distant tissues.
Autonomic & Structural Efferents (Orange pathways): Interoception triggers continuous, top-down physiological adjustments via the autonomic nervous system. The vagus nerve acts as a massive bidirectional conduit; after receiving sensory data, it sends parasympathetic efferent commands (routed back through the NTS and adjacent motor nuclei) down to the gut to regulate motility and secretion. Concurrently, sympathetic tracts dynamically regulate heart rate and blood pressure. Strikingly, the brain’s hardwiring extends deep into structural anatomy; direct sympathetic projections innervate the spleen and the marrow of bone, allowing the brain to physically communicate with the stem cell niche to regulate daily immune cell production.
A brain is a control system, and a control system is only as good as its connection to the thing it controls. A thermostat wired to nothing regulates nothing. The brain spends roughly a fifth of your entire resting energy budget — a staggering overhead for an organ that is about 2% of your body mass — and evolution does not subsidize expensive organs out of generosity. Recall the framing from Chapter 1: the costly brain earns its keep by buying prediction, by forecasting a need before the error arrives rather than merely reacting once it has. But you cannot predict what you cannot sense, and you cannot regulate what you cannot reach. So before the brain can be a predictive controller of the body, it has to be wired into the body — densely, bidirectionally, with dedicated channels carrying information up and commands down.
That wiring is the subject of this chapter. The peripheral nerves, the vagus nerve, the blood supply, and its barriers and gaps are not merely infrastructure. They are the sensory and motor apparatus of a control system whose controlled variable is the internal state of the body itself. The brain is not a disembodied processor that happens to sit on top of a body and issue orders. It is a node inside a set of feedback loops that run out through the body and back. Most of what the brain regulates, it regulates through these loops, by sensing the body’s internal condition, integrating those signals, and acting on them.
This chapter is therefore a hinge. It is not yet the full anatomy tour, which comes in Chapter 5. It is also not the sensory-systems unit, where we will fully explicate vision, audition, touch, taste, smell, and the cortical machinery that builds a world from them. Nor is it the detailed look at the hypothalamus (Chapter 6), where we will consider the regulatory machinery in detail. The goal here is simpler and more foundational: to show how the brain is coupled to the body it regulates. The nervous system has a peripheral half; the body sends signals upward through a variety of mechanisms; the brain sends signals downward; and the loop itself is the unit of regulation.
There is a word for the sensing half of this—the perception of the body’s own internal physiological state. It is interoception, and it underlies this whole chapter [@craig2002; @critchley2013]. Hunger, thirst, nausea, breathlessness, the racing heart, the full bladder, the warmth of a fever, the malaise of an infection—these are not incidental feelings layered on top of cognition. They are reports from the viscera, and the brain’s reading of them is the substrate on which allostatic regulation is built [@sterling2012]. When Peter Sterling argues that the brain’s basic job is predictive regulation of the body, the interoceptive channels provide the data it needs to make those predictions.
It is tempting to describe all this by casting the brain as a little commander “reading dispatches” and “issuing orders.” If you find me using such descriptions, it is only because that language is vivid. There is no homunculus sitting in the brain’s situation room. There are loops—afferent fibers carrying signals from body to brain, efferent fibers carrying signals from brain to body—and the regulation is the loop, not a decision made above it.
This chapter uses directional language, so here is the minimum vocabulary. Refer back to this list when a direction word trips you up.
- Direction of flow: Afferent means carrying signals toward the central nervous system (body → brain); efferent means carrying them away from it (brain → body). A useful mnemonic: Afferent = Arriving, Efferent = Exiting.
- Relative to the midline: Medial means toward the midline or center, and lateral means toward the outside or edge. Ipsilateral refers to structures on the same side of the body, while contralateral refers to structures on opposite sides.
- Front to back: Anterior (front) and posterior (back) are often used interchangeably with rostral (toward the beak/nose) and caudal (toward the tail).
- Top to bottom: Superior (above) and inferior (below) describe vertical positions. In neuroanatomy, we also use dorsal (toward the back) and ventral (toward the belly). Because humans walk upright, our neuraxis bends at the neck; therefore, dorsal points up (superior) in the brain, but points backward (posterior) in the spinal cord.
- Planes of section: When viewing brain slices or scans, the sagittal plane divides the left and right sides, the coronal plane divides the front and back (like a crown), and the axial (or horizontal) plane divides the top and bottom (parallel to the floor).
- The spinal division: In the spinal cord, the dorsal side is sensory (signals arriving) and the ventral side is motor (signals exiting)—a division that, as we saw last chapter, traces straight back to the alar and basal plates of the neural tube.
- The gross plan, in one line: The embryonic neural tube swells into three primary vesicles (forebrain, midbrain, hindbrain), which subdivide into five, which mature into the adult structures we will tour in the next chapter. For now, two landmarks matter: the hindbrain, where much of the body’s sensory traffic first lands, and the hypothalamus (in the forebrain), where much of the body’s regulation is orchestrated.
4.1 The nervous system has two major components
We often say “the brain” and “the nervous system” as if they were the same thing. They are not. The central nervous system (CNS) is the brain and spinal cord — the part encased in bone. The peripheral nervous system (PNS) is everything else: the nerves threading out to muscles, organs, skin, and gut. You cannot understand what the central nervous system is for without the peripheral nervous system, because the PNS is the interface. It is where the control system touches the controlled.
The PNS divides into two arms, and the division is functional. The somatic nervous system handles the “external” relationship with the world: voluntary control of skeletal muscle (efferent) and sensation from the skin, joints, and muscles (afferent). When you decide to lift a cup, the command travels out through somatic motor fibers whose cell bodies sit in the ventral horn of the spinal cord, exit through the ventral roots, and innervate the muscle. When you feel the cup’s warmth, the signal travels in through somatic sensory fibers whose cell bodies sit in the dorsal root ganglia — ganglia are clusters of neuronal cell bodies just outside the spinal cord — and enter through the dorsal roots. Sensory in the back (dorsal, or posterior), motor in the belly (ventral, or anterior). This corresponds to the alar/basal plate as discussed in Chapter 3.
That somatic loop is very important, but we will defer a detailed discussion of this system until Chapter 21. Later, when we study touch, pain, vision, hearing, smell, taste, and balance, we will ask how nervous systems build a model of the outside world. Here, and in Chapter 6, we will discuss the other half of the story: how the nervous system senses and regulates the body from within.
Open the interactive diagram in a new tab ↗
The autonomic nervous system (ANS) handles that “internal” relationship — the regulation of the viscera. This is the arm that matters most for our thesis, so it gets most of this section. But notice the shape of the whole thing first: the somatic system points outward at the world, the autonomic system points inward at the body, and both are loops with an afferent leg and an efferent leg. The brain’s relationship to your hand and the brain’s relationship to your heart are built from the same basic parts.
4.1.1 The autonomic nervous system, beyond “fight or flight”
Almost every introductory account of the autonomic nervous system reaches immediately for “fight or flight,” and I want to start by gently prying that phrase loose, because this pithy phrase has the power mislead students.
Here is the standard picture, which is not wrong, just poorly framed. The ANS has two major divisions that innervate the same organs with generally opposing effects. The sympathetic division is the accelerator: it speeds the heart, dilates the pupils and the airways, mobilizes glucose, and shunts blood toward muscle — the body configured for exertion. The parasympathetic division is the brake: it slows the heart, constricts the pupil, and promotes digestion — the body configured for maintenance. “Fight or flight” versus “Rest and digest.” So far, so textbook.
The trouble is that this reduces the autonomic nervous system to an emergency system — something that switches on when a tiger appears and otherwise idles. That is almost the opposite of the truth. The autonomic nervous system is running right now, continuously, in both of us, with no tiger anywhere. It holds your blood pressure within a narrow band as you stand and sit, trims your heart rate to your metabolic demand breath by breath, manages your core temperature, your gut motility, your bladder, your blood glucose. The “opposition” of sympathetic and parasympathetic is not a toggle between calm and panic; it is the continuous push-pull of a regulator holding a controlled variable steady — and, when the system is working in allostatic rather than merely reactive mode, adjusting the set-point in anticipation of demand. Your heart rate begins to climb fractionally before you stand up, not after your blood pressure has already dropped. That is prediction, implemented in autonomic tone.
And the two divisions are not a simple seesaw. Plenty of coordinated behaviors require both arms active at once. Sexual arousal is the classic example — erection is largely parasympathetic, emission largely sympathetic — and you cannot describe it as either “fight or flight” or “rest and digest” without the description falling apart. The accelerator-and-brake metaphor is a fine, first handhold, but a real driver uses both pedals together more often than you might think.
It is easy to picture the autonomic nervous system as purely efferent — orders going out to organs. That picture is half the system. The viscera report back constantly: baroreceptors in your arteries reporting blood pressure, chemoreceptors reporting blood gases, stretch receptors in the gut and bladder reporting fullness. Most of this never reaches consciousness, which is exactly why it is easy to forget it is there. But it is the afferent leg of the loop, and without it the efferent commands would be flying blind. Keep both legs in view. We are about to meet a single nerve that carries the body’s afferent traffic in extraordinary volume.
4.1.2 The enteric nervous system
Before we get to that nerve, one autonomic subsystem deserves consideration. Lining the wall of your gut, from esophagus to rectum, is the enteric nervous system (ENS) — a mesh of approximately 400 to 600 million neurons, more than the spinal cord contains, organized into two interconnected plexuses [@furness2012]. It is sometimes called the “second brain,” and for once the nickname is defensible: the enteric nervous system can run the basic program of digestion — sensing the contents of the gut, coordinating the muscular waves of peristalsis, regulating secretion — without instruction from the brain at all. Sever its connection to the CNS and the gut keeps digesting. No other organ system has anything like this degree of local neural autonomy [@spencer2020].
For our purposes, the enteric nervous system teaches a limited but important lesson. The gut is not a passive bag of nutrients waiting for brain commands. It has local neural machinery, and that machinery is linked to the brain through autonomic, hormonal, immune, and sensory channels. The gut is therefore one of the clearest examples of the embodied brain: local regulation in the periphery, central regulation from the brain, and continuous communication between them.
The enteric nervous system is also the locus of many overhyped claims in popular culture.
A fact students enjoy, and which is true: a large majority of the body’s serotonin (often quoted as ~90%) and a substantial fraction of its dopamine are found in the gut, not the brain. This is real. What it does not license is the leap you will see in popular writing — that because “the happy chemical” is mostly in your gut, your gut controls your mood. Gut serotonin and brain serotonin are, for most purposes, separate pools; serotonin does not freely cross the blood–brain barrier, and enteric serotonin is mostly doing local jobs — driving motility, signaling to sensory fibers — that have nothing to do with how you feel on a Tuesday. The fact is true but the inference is mostly false. Indeed, the gut–brain literature is rife with true facts attached to overreaching inferences.
What is solid. The gut and the brain communicate, bidirectionally, through several channels — chiefly the vagus nerve, but also hormonal and immune signals. Enteric sensory cells, including specialized enteroendocrine cells in the gut lining, detect the chemical state of the gut’s contents and relay that information to the brain. This much is not in doubt.
What is suggestive but unsettled. The claim that the community of microbes living in your gut — the microbiota — influences brain function and behavior. There is, at this point, a genuinely large body of evidence that the gut microbiota and the brain are connected [@cryan2019; @morais2021]. In rodents, the evidence for causation is in places quite strong: germ-free mice raised with no microbiota at all show altered stress responses and altered brain development, and — this is the striking one — transferring the gut microbiota from a depressed human into a microbiota-free rodent can transfer depressive-like behavior to the animal. That is a causal experiment, and it points to something real.
What is genuinely uncertain. Whether any of this scales to human mood, cognition, or psychiatric disease in the strong way the supplement aisle at CVS implies. The honest summary, and here I am leaning on a pointed review by Jenn Walter and her colleagues with a title I admire — “Establishing or exaggerating causality for the gut microbiome” — is that the human evidence is overwhelmingly correlational [@walter2020]. We see that the microbiota differs between depressed and non-depressed people, between people with and without various conditions. We do not, for the most part, know which way the arrow points. Does dysbiosis contribute to depression, or does depression — via diet, stress, and altered gut physiology — reshape the microbiota? Cross-sectional differences cannot tell us, and the rodent models, for all their power, translate to humans poorly and inconsistently. Probiotic trials in humans have shown, at best, modest antidepressant effects, and the field has a replication problem it is still working through.
My own view is this: the microbiota–gut–brain axis is real, it is important, but it is one of the most oversold topics in contemporary neuroscience. Both halves of that sentence are true at once. Be excited and be skeptical.
Here is a case that shows both how seriously to take gut–brain communication and how carefully to state what we know.
There is a hypothesis, originally from neuropathologist Heiko Braak, that some cases of Parkinson’s disease may begin in the gut and ascend to the brain along the vagus nerve [@braak2003]. The reasoning starts from an autopsy observation: the misfolded protein that defines Parkinson’s pathology, α-synuclein, often appears early in the dorsal motor nucleus of the vagus and in the nerve plexuses of the gut — earlier, in some patients, than in the substantia nigra, the midbrain region whose dopamine-neuron loss causes the disease’s motor symptoms. Braak proposed that pathological α-synuclein might propagate, prion-like, from the enteric nervous system up the vagus to the brainstem and onward.
For years this was an intriguing pattern in postmortem tissue and not much more. Then, in 2019, Kim and colleagues did the experiment in mice: they injected pathological α-synuclein into the gut wall and watched it spread — first to the dorsal motor nucleus of the vagus, then up through the brainstem, and eventually to the substantia nigra, accompanied by dopamine-neuron loss and Parkinson-like motor and non-motor deficits. Crucially, cutting the vagus nerve (truncal vagotomy) blocked the spread [@kim2019]. And there is supporting human epidemiology: people who underwent truncal vagotomy for ulcer disease decades ago appear to have a somewhat reduced later risk of Parkinson’s.
This is a striking convergence of pathology, a causal animal model, and human epidemiology, all pointing along the anatomical route this chapter is about. So why am I filing it under “caution” rather than “established”? Because the strong claim — that Parkinson’s is, in general, a gut-origin disease — remains unproven in humans [@horsager2020]. The propagation looks bidirectional: there appear to be “body-first” cases that fit Braak’s gut-to-brain route and “brain-first” cases that do not, and we cannot yet say what fraction of disease each accounts for. The vagus is clearly a highway; whether it is the origin is unsettled.
Be wary of the version of this you may encounter in popular health writing, which flattens “Parkinson’s may have a body-first subtype that involves the vagus” into “Parkinson’s starts in your gut, so fix your gut.” The careful claim is exciting enough. It does not need the inflation. You will also see the gut floated as the origin of ALS and autism. The evidence there is far thinner than for Parkinson’s, and I would not lump them together; the Parkinson’s case is in a different evidential class.
Because the autonomic nervous system drives peripheral organs in measurable ways, we can read out a person’s autonomic state from the outside, non-invasively. This is the basis of psychophysiology, and it is worth knowing both because the measures recur throughout the course and because they make the abstract idea of “autonomic tone” concrete.
The general logic is that sympathetic activation produces a family of bodily changes — and we can instrument each of them. The common measures:
- Pupillometry. The pupil dilates with arousal, attention, and cognitive load, and constricts with parasympathetic dominance. Tracking pupil diameter gives a surprisingly sensitive, fast index of mental effort.
- Electrodermal activity — the skin conductance response (SCR), historically the galvanic skin response (GSR). Sweat glands are under sympathetic control; sweaty skin conducts a small electrical current better than dry skin. A spike in skin conductance is a fairly direct readout of a sympathetic burst.
- Heart rate and heart rate variability (HRV). Heart rate rises with sympathetic drive; variability between successive beats tends to fall under stress and rise during relaxation. HRV in particular is heavily shaped by parasympathetic (vagal) influence on the heart, which is one reason it has become a popular — sometimes over-interpreted — index of “vagal tone.”
- Respiration. Breathing rate and depth shift with arousal.
You have met this technology in one well known and scientifically dubious application: the polygraph “lie detector” is essentially a multi-channel psychophysiology rig — skin conductance, heart rate, respiration — recording arousal during questioning. The reason polygraphy is unreliable is instructive and worth stating plainly: these measures index arousal, not deception. An innocent person terrified of a false accusation and a guilty person calmly lying can produce overlapping traces, because the autonomic nervous system does not know or care whether you are telling the truth — it responds to how threatened you feel. The instruments are real and useful; the inference from arousal to lying is the weak link. Notice that this is the same lesson as the serotonin example and the microbiome example: a real signal, a tempting inference, and a gap between them. I promise I am not arranging these on purpose. The gap just keeps showing up, which is itself the point.
4.2 The vagus nerve
The vagus nerve is the tenth cranial nerve. Its name comes from the Latin for “wandering,” and it earns it: from its origins in the medulla it wanders down through the neck and thorax into the abdomen, branching to innervate the larynx, heart, lungs, and most of the gastrointestinal tract. It is the longest and most widely distributed of the cranial nerves, and it is the principal conduit of the parasympathetic nervous system. So far this sounds like an efferent story — the brain’s long arm reaching down to slow the heart and stimulate digestion. And the vagus does carry those commands.
But here is the fact that reorganizes the whole picture. The vagus is overwhelmingly a sensory nerve. Roughly 80% of its fibers — the figure is sometimes given as 80 to 90%, and the exact number has a long pedigree in the physiology literature — are afferent, carrying information from the body up to the brain [@berthoud2000; @prescott2022]. Only about 20% are efferent. The nerve we describe as the brain’s great parasympathetic output is, in the brute accounting of its fibers, mostly an input. The body reports up far more than the brain commands down.
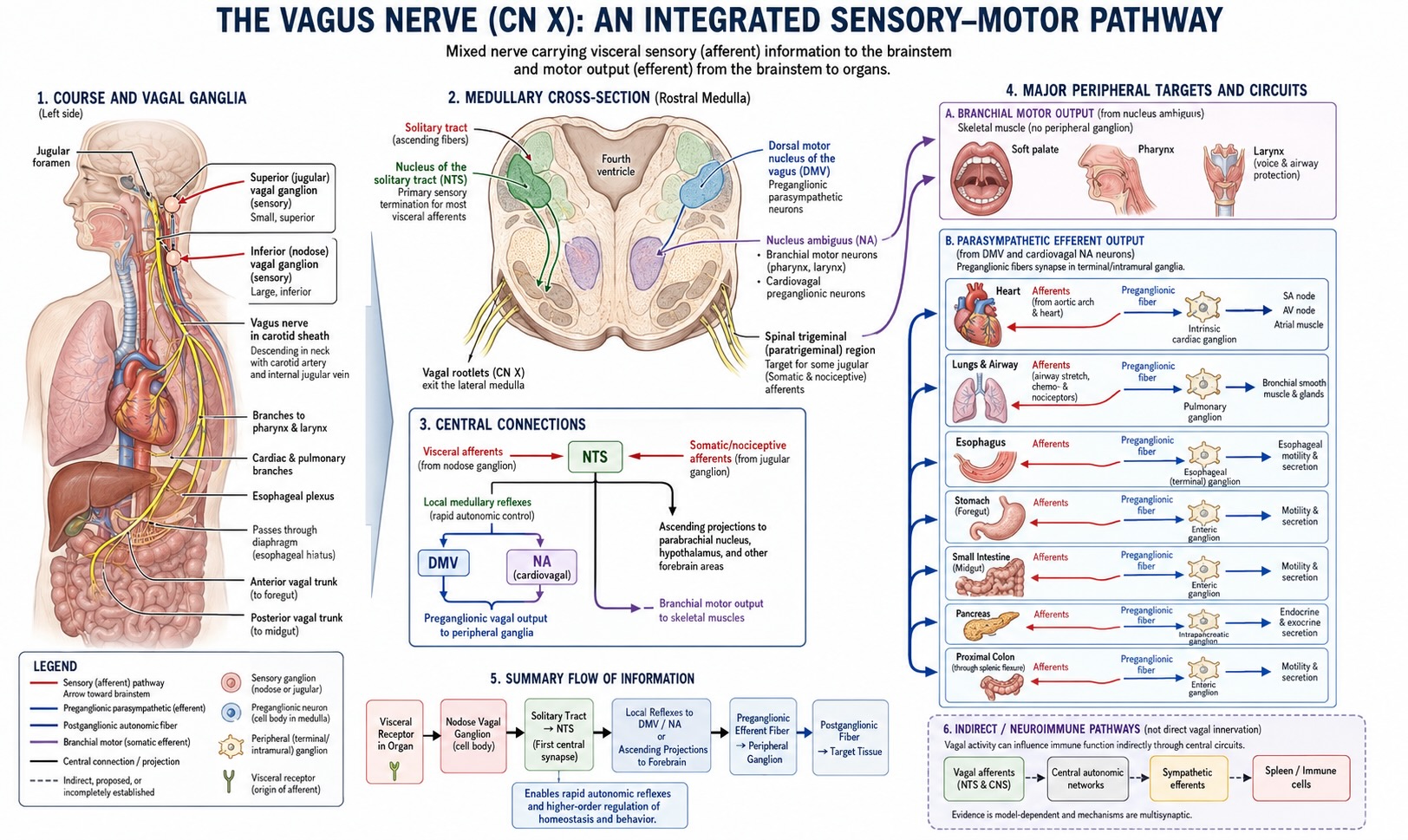
This inverts the intuitive model. We imagine the brain as the body’s commander, issuing a high volume of orders to a passive periphery. The vagus says the reverse: the dominant flow of traffic, by a factor of four, is the periphery reporting its state to a brain whose main job is to listen and adjust. The heart, the lungs, the gut, the great vessels — all of them are continuously streaming status reports upward: blood pressure, oxygen and carbon dioxide levels, gut distension, the chemical contents of a meal, the presence of inflammation. The brain is, more than anything, an organ that is being constantly informed about the body. This is interoception with an anatomical address.
Where does all this afferent traffic land? Overwhelmingly at one structure: the nucleus of the solitary tract (NTS; also called the nucleus tractus solitarii), a column of cells in the medulla. About 80% of vagal afferent fibers terminate here, making the NTS the brain’s primary visceral-sensory relay — the great clearinghouse where the body’s reports first arrive in the central nervous system [@saper2002]. It is the visceral analogue of the way the thalamus relays the “external” senses, and it is far less famous than it deserves to be.
The NTS does not keep this information; it distributes it, and the projection targets are exactly the structures you would design a regulatory system to inform. It projects to the parabrachial nucleus (a major waystation in the pons that integrates visceral and other signals), to the hypothalamus — the master regulator of homeostasis, which we will return to repeatedly — and onward to limbic and forebrain regions involved in the feeling and the behavioral consequences of bodily states [@saper2002]. So the path is: body → vagal afferents → NTS → parabrachial nucleus / hypothalamus → integration and response. That is the afferent half of the loop, drawn end to end, and the hypothalamus sitting at its terminus is not an accident. The body’s status reports flow to the structure whose job is to keep the body in balance.
The important point for now is not to memorize the NTS, parabrachial nucleus, and hypothalamus as disconnected names. The important point is that the body has a main sensory entrance into the brainstem, and that entrance points toward the regulatory machinery. In Chapter 5, when we tour the CNS, and in Chapter 6, where we discuss the hypothalamus, the brainstem will no longer be a stalk underneath the brain. It will be the first central station for the body’s reports.
4.3 The NTS is evolutionarily old
In keeping with this book’s emphasis on evolution, it should not be surprising that the NTS has a long history. It is a deeply conserved structure with recognizable homologues across the vertebrates [@nts_vertebrates2026]. The lamprey has one; the fish has a prominent one. In some fish the vagal and facial sensory lobes are among the largest structures in the brain, because taste and visceral sensation matter enormously to an animal feeding in water. A bird’s NTS contains a topographic map of the viscera.
This is what we should expect. Sensing the internal state of the body and adjusting physiology to match is not a luxury that arrived with mammalian sophistication; it is among the oldest jobs a vertebrate brain has, because it is among the most necessary. An animal that cannot monitor and regulate its own viscera does not live long enough to evolve anything fancier. The elaborate human forebrain is a late and expensive addition built on top of this ancient body-regulating core — not a replacement for it. We are, here as elsewhere, a modified vertebrate.
4.4 Breaching the barrier: the circumventricular organs
The vagus is the brain’s neural line to the body — fast, wired, point-to-point. But the body also speaks to the brain in a slower, broadcast medium: the bloodstream, carrying hormones, metabolites, and immune signals. And here we hit an apparent problem, one that sets up the next two sections.
The brain, almost everywhere, walls itself off from many molecules and pathogens carried in the blood. The blood–brain barrier is the name for this: the endothelial cells lining the brain’s capillaries are stitched together with tight junctions, and unlike the leaky capillaries elsewhere in the body, they do not let molecules slip freely from blood into brain tissue [@daneman2015]. This is, on the whole, a feature and not a bug — the brain is a delicate electrochemical instrument, and you do not want every fluctuation in blood chemistry, every ingested toxin, every circulating hormone to directly affect your neurons. The barrier keeps the brain’s internal environment tightly controlled.
But now consider the regulatory problem this creates. If the brain is supposed to keep the body in homeostasis, it needs to know the body’s blood chemistry — the osmolality (saltiness) of the blood, its glucose, its hormone levels, whether there are toxins or inflammatory signals present. How can the master regulator monitor the blood if it has deliberately walled itself off from the blood?
The answer is elegant: the brain leaves itself a set of windows. The circumventricular organs (CVOs) are small midline structures bordering the ventricles where the blood–brain barrier is deliberately absent — the capillaries there are fenestrated (leaky, windowed) rather than sealed [@miyata2015; @mckinley2003]. At these specific, limited sites, and essentially nowhere else, neurons are directly exposed to the chemical environment of the blood. They are sampling ports — places where the brain pokes a sensor through its own barrier to sample the blood directly.
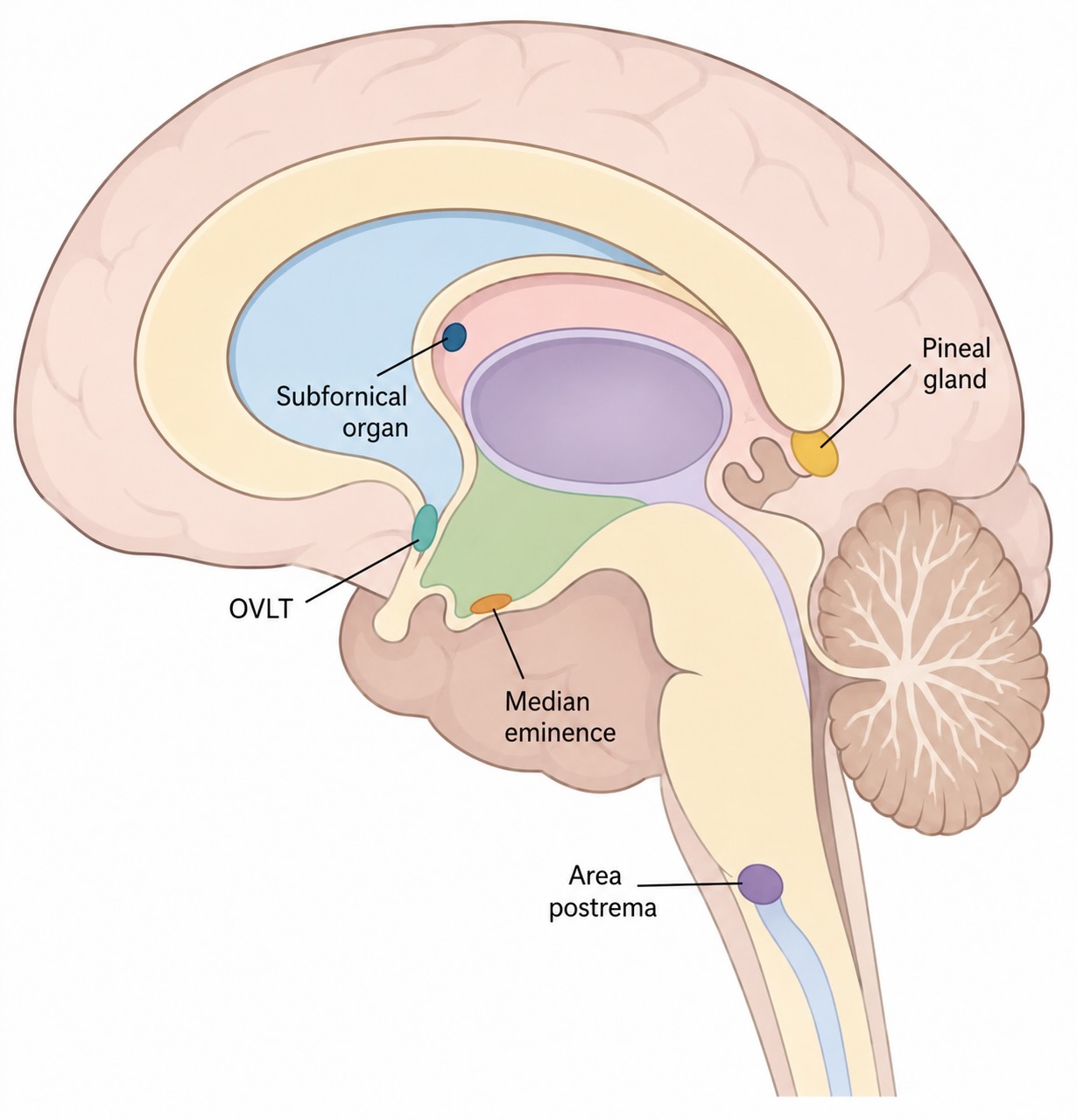
It helps to split the CVOs into two functional groups, because they face in opposite directions:
- Sensory CVOs — chiefly the subfornical organ, the organum vasculosum of the lamina terminalis (OVLT), and the area postrema. These contain neurons exposed to the blood that sense its contents and project that information inward to the rest of the brain. The subfornical organ and OVLT monitor blood osmolality and circulating signals related to fluid balance and blood pressure — they are central to how the brain knows you are dehydrated and generates thirst. The area postrema, in the floor of the fourth ventricle, is a chemoreceptor trigger zone: it samples the blood for circulating toxins, and when it detects them it can trigger vomiting. This is why the area postrema is sometimes called the “vomiting center,” and why a number of drugs cause nausea as a side effect — they are being detected at this deliberately unguarded window. Tellingly, the area postrema projects directly to the NTS, projecting the humoral sensing of the CVOs into the same visceral-relay machinery the vagus feeds. The two channels — neural and bloodborne — converge.
- Secretory CVOs — chiefly the median eminence and the neurohypophysis (posterior pituitary), plus the pineal gland. These face the other way: rather than sensing the blood, they secrete into it. The median eminence is where hypothalamic releasing hormones are dumped into the special portal blood supply that carries them to the pituitary; the neurohypophysis releases oxytocin and vasopressin into the general circulation; the pineal secretes melatonin. These are the brain’s output ports to the bloodstream.
So the circumventricular organs are the missing mechanistic link between two ideas that the room-by-room anatomy tour leaves disconnected: the brain’s “plumbing” and the brain’s sensing. The blood is not just delivering fuel to the brain; at these windows, the brain is reading the blood as a sensory surface. And once you see that the brain has dedicated machinery for sampling bloodborne signals, the door opens to a question the older textbooks barely asked: what is in the blood that the brain wants to read? Hormones and salt and glucose, yes. But also the molecular signals of the immune system.
This is another place where we are deliberately previewing future units rather than teaching them fully. Thirst, salt appetite, hunger, fever, stress hormones, circadian rhythms, and reproductive hormones all pass through hypothalamic and endocrine machinery that we will return to later. For now, CVOs give you the architectural fact: the brain has windows through which it samples the body directly.
4.5 The vascular system: supply and signal
We have arrived, by the logic of sensing, at the brain’s blood supply — and I want to treat it in a way that the usual “here is the Circle of Willis, memorize the arteries” presentation does not. The vascular system has three jobs that we discuss in below. The first is obvious while the other two are not.
4.5.1 The first job: fuel
The first job is the obvious one. The brain runs on glucose and oxygen, both delivered by blood, and it runs on them with almost no reserve. Recall the budget from earlier chapters: the brain consumes roughly 20% of the body’s resting energy — on the order of several hundred kilocalories a day just to keep ~86 billion neurons supplied. Herculano-Houzel’s figure of about 516 kcal/day for the human complement of neurons is the number we have been carrying. Now add a crucial fact: unlike muscle, which stockpiles energy as glycogen, the brain stores almost none (and most of what it stores is found in glia cells, not neurons). It is a high-draw device utterly dependent on continuous delivery of energy.
This is why interruption of blood flow to the brain is catastrophic on a timescale of seconds. Cut the supply and consciousness is lost within seconds; let the interruption persist and the deprived tissue dies. This is a stroke. In an ischemic stroke, the common kind, an embolus — typically a fragment of atherosclerotic plaque — travels along an artery until it lodges where the vessel narrows too far for it to pass, blocking flow to everything downstream. The tissue served by that vessel is starved of glucose and oxygen and becomes infarcted: it dies, and the functions it supported are lost — a paralysis, a speech deficit, a field of blindness, depending on what the blocked artery fed.
Stroke is a tragedy for the patient and, historically, one of the most important tools neuroscience has had. Because a stroke knocks out a specific, vascularly-defined territory and produces a specific loss of function, the correspondence between the two has been a primary source of our knowledge of what brain regions do. Much of classical localization — including the language regions we will study later — was first charted by correlating the site of a patient’s lesion, often a stroke, with the function they lost. The brain’s blood supply is, among other things, a map of how the brain can fail, and how it fails has taught us how it works.
The supply itself, in brief: blood reaches the brain by two routes. The paired internal carotid arteries ascend through the neck and feed mainly the anterior brain — the “anterior circulation.” The paired vertebral arteries ascend along the spine and fuse into the single basilar artery, supplying the brainstem, cerebellum, and the posterior brain including the occipital lobe — the “posterior circulation.” These two systems meet in a ring at the base of the brain, the Circle of Willis, from which the major cerebral arteries branch. The ring is a piece of engineering redundancy: in principle, if one feeder is compromised, the ring allows flow to cross over and compensate. In practice the compensation is partial and varies between individuals, but the design intent — a shared reservoir rather than independent end-arteries — is clear.
4.5.2 The second job: signal
Now the part that earns the vascular system its place in this chapter rather than the next. The blood is not only a fuel line. It is a communication channel, and the brain both reads it and writes to it.
We have already seen the reading machinery — the circumventricular organs, where the brain samples circulating hormones, osmolality, toxins, and immune signals. And we have seen the writing machinery — the secretory CVOs, where the brain dumps hormones into the blood to act at a distance. Put those together and the vascular system becomes a third great input/output system of the brain, parallel to the neural one. The vagus and the spinal nerves are the brain’s wired connection to the body — fast, specific, addressed. The bloodstream is the brain’s wireless connection — slower, diffuse, broadcast. Both carry the body’s state up and the brain’s regulation down. A control system wired into its body by only nerves would be missing half the conversation; the body’s chemistry — its hormones, its fuels, its inflammatory state — is carried in blood, and the brain has evolved the apparatus to participate in that chemical conversation in both directions.
4.5.3 The third job: clearance
The brain’s blood vessels help organize the clearance of waste. Every active tissue makes trash. Neurons and glia generate ions, neurotransmitter breakdown products, misfolded proteins, inflammatory molecules, and ordinary metabolic by-products that cannot simply accumulate in the extracellular space. This third job of the vascular system intersects with our discussion of the brain’s glymphatic and lymphatic systems that will be discussed in detail in Section 4.7.
There is a beautiful consequence of the brain’s continuous need for energy and it provides a means by which we can image the living, working human brain at all.
When a patch of brain becomes more active, it demands more fuel, and local blood flow to that patch increases to meet the demand — a phenomenon called functional hyperemia, first noted by Roy and Sherrington in 1890 [@roy1890]. The local matching of blood flow to neural activity is called neurovascular coupling, and it is mediated by a little consortium of cells — neurons, astrocytes, and the vessels themselves — sometimes called the neurovascular unit [@iadecola2017]. The upshot is that where the brain is working, the blood follows. And that means if you can image local blood flow or blood oxygenation, you can infer where neural activity is occurring.
This is exactly what the two great functional-imaging methods do. PET (positron emission tomography) and, more importantly today, functional MRI (fMRI) both work by, in effect, following the energy — fMRI by detecting the blood-oxygenation changes, the “BOLD” signal, that accompany the hemodynamic response to neural activity. When you read that a study “showed activation” in some brain region, what was almost always measured was not neural firing directly but the blood-flow change that neurovascular coupling produces a couple of seconds later. We will take this up in detail when we cover imaging methods. For now, notice the double duty the vascular system is doing in this chapter: it is both an instance of body–brain signaling and the tool that lets us watch the brain signal.
A more detailed look at the methods used in human neuroscience research can be found in Appendix B.
4.6 The immune–brain axis
For most of the twentieth century, the brain was considered immune-privileged — walled off from the immune system, sitting behind the blood–brain barrier, and lacking lymphatic drainage. The dogma had real evidence behind it. It was also, we now know, substantially wrong, or at least far too strong. The brain and the immune system are in constant communication, in both directions. Let me start with the part that is not speculative.
4.6.1 The solid anchor: sickness behavior
Start with something you have experienced. When you come down with the flu, you do not merely have a fever and aches. Your behavior changes, in a characteristic and coordinated way: you lose your appetite, you withdraw from social contact, you become sleepy and listless, you lose interest in things that normally motivate you. We tend to think of this as just “feeling bad” — an unfortunate by-product of being sick. It is not a by-product. It is a program, organized by the brain, and it has a name: sickness behavior [@hart1988; @dantzer2008].
Here is the chain of events, which is well established. When you are infected, cells of your innate immune system release signaling molecules called pro-inflammatory cytokines — interleukin-1β, TNF-α, interleukin-6, and others. These cytokines are the immune system’s local messengers, coordinating the inflammatory response in the body. But the brain detects them, and in response it orchestrates the suite of sickness behaviors [@konsman2002]. The lethargy, the anorexia, the social withdrawal, the fever itself — these are brain-driven outputs, triggered by the brain’s reading of immune signals.
Sickness behavior is adaptive. It is not the infection disabling you; it is your brain reallocating your resources to fight the infection. Fever is metabolically expensive and is run deliberately because many pathogens replicate poorly in high temperatures. Anorexia and lethargy conserve the energy that fever and immune activation demand, and reduce your exposure to new pathogens and predators while you are compromised. Social withdrawal may reduce transmission to kin. The whole package is a coordinated, evolved, brain-mediated shift in physiology and behavior in the service of survival. Which is to say: sickness behavior is allostasis — predictive reallocation of the body’s resources in anticipation of need — implemented in response to an immune challenge. The immune system reports a threat; the brain reconfigures the entire organism to meet it.
Now, how do the cytokines reach the brain? This is where the routes appear, and notice that the first two are channels we have already built in this chapter:
- The neural route. Vagal afferents detect cytokines in the body’s tissues and relay the inflammatory signal up to the NTS — the same fast, wired pathway we traced earlier. This is the rapid arm: the brain learns of peripheral inflammation through its sensory nerves, the same way it learns of a full stomach or a drop in blood pressure. It is interesting to note that the vagus does not only report inflammation upward; through its efferent fibers it can suppress inflammation downward, via what Kevin Tracey named the cholinergic anti-inflammatory pathway or “inflammatory reflex” [@tracey2002; @borovikova2000]. Efferent vagal activity, through acetylcholine, can dampen the production of inflammatory cytokines by immune cells in the body. So the vagus is a true two-way regulatory loop for the immune system specifically: afferents sense peripheral inflammation, the brainstem integrates it, and efferents can turn the inflammation down.
- The humoral route. Cytokines circulating in the blood act at the circumventricular organs — the windows in the barrier we just met — and drive the local production of further signals, such as prostaglandins, that propagate the inflammatory message into the brain proper. This is the slower arm, and it is exactly the “reading the blood as a sensory surface” idea from the CVO section, now with immune molecules as the thing being read.
So sickness behavior is not some exotic new system. It is the afferent/efferent machinery of this entire chapter — vagus, NTS, CVOs, hypothalamus — being used to sense and respond to the immune system. That is why I can call it solid: the routes were independently established, the cytokines are identified, the behavioral program is reproducible, and the adaptive logic is coherent.
If acute immune signaling to the brain produces adaptive sickness behavior, what happens when immune signaling becomes chronic — in persistent infection, autoimmune disease, chronic stress, or the low-grade inflammation associated with obesity and aging?
The hypothesis, associated especially with Dantzer and colleagues, is that sustained inflammatory signaling to the brain can tip sickness behavior over into something that looks like clinical depression — and that inflammation may therefore be one contributing pathway to depression in some people [@dantzer2008]. The overlap between sickness behavior and the symptoms of depression — anhedonia, fatigue, social withdrawal, appetite and sleep disruption — is hard to ignore. There is supporting evidence: patients given inflammatory cytokines as therapy, such as interferon-α for hepatitis or cancer, develop depression at strikingly high rates, and meta-analyses find elevated inflammatory markers in depressed populations.
I find this genuinely persuasive as one pathway among several. Inflammation is not “the cause of depression”; depression is heterogeneous, and most depressed people are not detectably inflamed. The defensible claim is that there appears to be an inflammation-linked route into depressive states in some individuals, which is both scientifically important. However, it is a long way from the headline “depression is an inflammatory disease.”
4.7 Brain drainage: glymphatic flow, meningeal lymphatics, and the skull
In most of the body, one part of the solution to the waste problem is the lymphatic system. Excess interstitial fluid and dissolved material enter lymphatic capillaries, pass through lymph nodes, and eventually return to the venous blood. The brain is different, but the difference needs to be stated carefully. The brain parenchyma — the tissue where neurons and glia live — does not contain the ordinary lymphatic capillary network found in many other organs. Yet the brain has an exceptionally high metabolic rate, so the clearance problem is especially acute.
One answer appears to be a vascular-adjacent clearance system. Cerebrospinal fluid can move along spaces surrounding penetrating arteries, exchange with interstitial fluid in the tissue, and help carry soluble waste products out along perivascular and perivenous routes. Because this pathway depends heavily on glial cells, especially astrocytic endfeet wrapped around blood vessels, and because it performs a lymph-like clearance function, it has been called the glymphatic system — “glial” plus “lymphatic” [@iliff2012; @xie2013; @bohr2022].
This does not mean that blood simply washes through the brain. The blood-brain barrier remains a barrier. The point is subtler and more interesting: blood vessels provide the physical tracks around which fluid movement is organized. Arterial pulsation, respiration, vascular dilation, and the state of sleep and wakefulness all appear to influence how fluid moves through these surrounding spaces. Recent animal work even suggests that neurovascular coupling — the increase in local blood flow that follows local neural activity — may help accelerate glymphatic influx and clearance in the activated region [@holsteinronsbo2023]. If that general idea holds, then increased blood flow is not only about delivering glucose and oxygen. It may also help maintain the chemical cleanliness of the extracellular space.
Where does the material ultimately go? Probably not to a single exit. Some solutes are handled through transport across the blood-brain barrier and other barrier systems. Some reach cerebrospinal-fluid spaces. Some drain along venous and perivascular routes. And some appears to leave through lymphatic vessels in the meninges, especially the dural lymphatics that drain toward the deep cervical lymph nodes [@louveau2015; @aspelund2015; @absinta2017].
This is why the older slogan “the brain has no lymphatic system” now needs a footnote. The brain tissue itself lacks ordinary lymphatic capillaries, but the membranes around the brain do contain genuine lymphatic vessels. These meningeal lymphatics were described convincingly in 2015 by Louveau and colleagues and by Aspelund and colleagues, although the story is also a rediscovery: earlier anatomical reports of lymphatic drainage around the brain had been forgotten, dismissed, or lacked the markers needed to settle the question [@louveau2015; @aspelund2015].
The current state of the field is best taught with two levels of confidence. The existence of meningeal lymphatic vessels is now well established, including in humans and non-human primates [@absinta2017]. Their function is also supported strongly in animal models: impairing meningeal lymphatics in mice slows macromolecular clearance, worsens some measures of cognitive performance, and can aggravate amyloid pathology in Alzheimer-model animals [@damesquita2018]. What remains less settled is the magnitude of their role in ordinary human brain clearance and in human disease. In Alzheimer’s disease, for example, meningeal-lymphatic dysfunction could be a contributing cause, a consequence, an aggravating factor, or sometimes a bystander. The anatomy is solid; the clinical weight of the anatomy is still being worked out.
The glymphatic model should be given the same balanced treatment. It is influential, experimentally productive, and supported by important animal and imaging evidence. It also remains more debated than the anatomy of meningeal lymphatics, especially when the question is the magnitude of bulk flow through human brain tissue under normal conditions. For this chapter, the safest conclusion is this: the brain is not a sealed, immune-isolated organ with no drains. It has barrier systems, perivascular clearance routes, cerebrospinal-fluid pathways, and meningeal lymphatics that connect the brain’s borders to the immune system’s filtration network.
That gives the vascular system a third job in this chapter. Blood vessels do not merely deliver fuel. They also provide routes for chemical signaling and help organize clearance. Fuel, signal, clearance — three vascular jobs, not one.
The skull is not just inert armor. Like other bones, it contains bone marrow, the tissue that produces blood cells, including immune cells. For a long time it was reasonable to assume that immune cells responding to brain injury or infection were recruited from the general circulation, with marrow throughout the body contributing more or less as a shared reservoir. Recent work has complicated that picture.
In mouse models of stroke and meningeal inflammation, Herisson and colleagues found microscopic vascular channels running through the inner skull, connecting skull marrow with the dura and the brain’s surface [@herisson2018]. In those models, myeloid cells such as neutrophils could move from adjacent skull marrow toward the inflamed brain border. Similar channels have been identified anatomically in human skull samples, although their functional importance in living humans is still uncertain.
The field has moved quickly. Skull and vertebral marrow now appear to act, at least in mice, as local immune reservoirs for the meninges and other central-nervous-system borders [@cugurra2021]. The traffic may also run in the other direction: cerebrospinal fluid can access skull marrow through dura-skull channels, and CSF-borne signals can influence marrow niches and myeloid-cell production after injury [@mazzitelli2022]. That gives the brain’s border a more reciprocal organization than older textbook diagrams suggested. Immune cells can be supplied locally from nearby marrow, and signals from the central nervous system can reach that marrow.
There is also a direct neural layer to the story. Bone marrow is innervated. Sympathetic fibers regulate hematopoietic stem and progenitor cell trafficking from marrow into the blood, partly through noradrenergic effects on stromal cells and chemokine signals in the marrow niche [@katayama2006; @mendezferrer2008]. Pain-sensing fibers can participate in marrow mobilization as well, at least under experimental conditions that stimulate hematopoietic stem-cell release [@gao2021]. The exact timing of these rhythms is species-specific — mouse and human mobilization rhythms are not simply aligned to the same clock phase — so the durable lesson is not a particular hour of the day, but the broader principle that marrow traffic is neurally and circadian regulated [@mendezferrer2008; @lucas2008].
I would not ask students to memorize this whole frontier. The organizing idea is the important part. The brain senses the immune system through cytokines, vagal afferents, circumventricular organs, and other body-interface routes. It also regulates immune physiology through autonomic output, vascular control, endocrine control, and now, perhaps, more local brain-border pathways involving meningeal lymphatics and skull marrow. The immune system is not a separate country with closed borders. It is another body system that the embodied brain senses, predicts, and regulates.
4.8 Coda: the machinery on which prediction runs
We started this unit with the claim that the brain is not a disembodied controller but a node in feedback loops that run out through the body and back. We have now seen the actual hardware of those loops.
The body reports its internal state upward through dedicated afferent channels — the vagus above all, carrying its afferent signals to the nucleus of the solitary tract; the spinal visceral afferents; the windows of the circumventricular organs where the brain samples blood chemistry directly; the cytokine signals of the immune system read through those same channels. These reports converge on integrating structures, the hypothalamus chief among them. And the brain acts back on the body through dedicated efferent channels — the sympathetic and parasympathetic arms of the autonomic nervous system continuously tuning the viscera, the hormonal outputs secreted into the blood, the cholinergic reflex restraining inflammation, the sympathetic fibers reaching into the bone marrow to clock the immune system’s output. Sense the body’s state; integrate it; act on it; sense the consequences. That loop, in all its parallel channels, is the embodied brain.
And now recall why this matters for the spine of the whole unit. The brain is expensive because it buys prediction — because allostasis, the anticipatory regulation of the body, is worth more than mere reactive homeostasis. But prediction requires data. You cannot forecast a need you cannot sense, and you cannot pre-empt an error in a variable you cannot measure. The afferent/efferent machinery of this chapter is precisely the apparatus that makes allostatic prediction possible: it is how the brain gets the continuous, high-bandwidth stream of bodily data that any predictive regulator must have, and how it reaches back to act before the error arrives. The interoceptive channels are not a side-system. They are the sensory and motor surface of the body-regulating control system that the expensive brain exists to be. The prediction the brain is buying has to run on something, and this is the something.
Which brings us, finally, to the structures themselves. We have spent this chapter on how the brain talks to the body — the channels in and out. We have deliberately deferred the brain’s internal organization: what the hypothalamus actually contains, how the cortex is arranged, what the cerebellum and basal ganglia and amygdala are doing, how the great fiber tracts wire the regions to each other. That is the subject of the next chapter, Chapter 5 — an anatomical tour of the human brain. Now that we understand the brain as an embodied controller — wired into the body it regulates — we are ready to tour the machinery that does the regulating, and to ask of each structure not merely “what is it called?” but “what is its role?”
In the spirit of the last two chapters, here is the ledger for this one.
What we’re sure of.
- The brain is densely, bidirectionally wired into the body. The afferent/efferent organization is not in doubt; the somatic and autonomic divisions and their basic anatomy are textbook-solid.
- The vagus is predominantly an afferent nerve (~80% of fibers) and its afferents land chiefly in the nucleus of the solitary tract, which relays visceral information to the parabrachial nucleus, hypothalamus, and beyond. Solid.
- The circumventricular organs are real windows in the blood–brain barrier where the brain samples, or secretes into, the blood. The sensory/secretory division and the major members are well established.
- The brain’s energy economy (≈20% of resting metabolism, negligible storage) and the consequent catastrophe of stroke are firmly established, as is neurovascular coupling as a phenomenon and the basis of fMRI.
- Sickness behavior is a real, adaptive, brain-organized response to immune signaling, communicated by well-characterized neural (vagal) and humoral (CVO) routes. Forty years of work; it holds.
What we’re not sure of (and where the hype lives).
- The microbiota–gut–brain axis is real, but enthusiasm is outstripping hard causal data. Rodent causal evidence is strong; human causal evidence is largely correlational, and the translation is poor. Be excited and skeptical at once.
- The Braak “gut-first” hypothesis for Parkinson’s has real support (a causal mouse model, vagotomy epidemiology) but is unproven as a general human account; the disease appears to have body-first and brain-first subtypes in unknown proportions.
- The mechanism of neurovascular coupling — astrocytes versus neurons; what the flow increase is even for — is genuinely debated, which matters because every fMRI result depends on it.
- The inflammation–depression link is persuasive as one pathway in some people, not as a theory of depression in general.
- The meningeal-lymphatic and skull-marrow-channel findings are real and confirmed to exist in humans, but most of the mechanistic and disease-relevance work is recent and rodent-derived. The organizing idea is more durable than any single finding; expect revision.