17 The cells of the brain
Neurons, Glia, Morphology, and Local Circuits
The brain is an organized tissue containing two broad cellular families: neurons and glia.
Neurons are electrically excitable cells specialized for signaling. They receive input, integrate it, and send output to other cells. Most neurons have recognizable input regions, an integrating cell body, and an output process called an axon. At the broadest functional level, the output of a neuron has two major effects on its target: it can excite the target, pushing it toward activity, or inhibit the target, pushing it away from activity. Later chapters will complicate that statement with neuromodulators, peptides, hormones, glial regulation, and all the biochemical messiness that makes real brains interesting. But the first grammar of neural signaling is excitation and inhibition.
Glia are the other major family of brain cells. The word comes from the Greek for “glue,” a historical name that is now mostly useful as a reminder of how badly these cells were underestimated. Glia do not usually send millisecond action potentials down axons, but they are not passive packing material. They regulate the chemical environment around neurons, insulate axons, defend and repair neural tissue, prune synapses, guide development, help form and maintain synapses, and help match local blood flow to local neural activity.
Unit 1 already introduced the headline number: the human brain contains roughly 86 billion neurons. The new fact for this chapter is that it contains roughly the same number of non-neuronal cells — about 85 billion — most of which are glia. The old textbook claim that glia outnumber neurons ten to one across the human brain is another example of a memorable number that survived longer than the evidence for it. The whole-brain ratio is closer to one, but that average hides enormous regional variation. The cerebral cortex has several non-neuronal cells for every neuron, while the cerebellum has very densely packed neurons and many fewer glia per neuron. A brain-wide number is a useful correction; it is not a substitute for anatomy.
This chapter introduces the cellular cast. We will begin with the major cell types, then examine the shape of neurons, the doctrine that made neurons the central unit of neuroscience, the developmental routes by which cortical neurons arrive in place, the ways neurons are classified, and the local circuit motifs that can be built from excitation and inhibition. We will then return to the glia and, especially, to astrocytes, which force us to stop imagining the synapse as a private conversation between two neurons.
17.1 The cellular cast, before the story begins
Here is the cast list before the narrative gets moving. Students should not have to hunt through a chapter to discover the major cell types in the brain.
| Broad class | Major types | What they do |
|---|---|---|
| Neurons | Excitatory neurons, inhibitory neurons, projection neurons, interneurons, modulatory neurons | Receive, integrate, and transmit signals; form local and long-range circuits |
| Astrocytes | Protoplasmic and fibrous astrocytes; specialized forms such as Bergmann glia and Müller glia in particular regions | Regulate extracellular ions and transmitters, support metabolism, wrap synapses, help form and modulate synapses, and contribute to neurovascular coupling |
| Oligodendrocytes | Myelinating glia of the central nervous system | Wrap CNS axons in myelin, increasing the speed and reliability of conduction |
| OPCs / NG2 glia | Oligodendrocyte precursor cells that persist in the adult brain | Generate oligodendrocytes, respond to injury, and participate in activity-dependent repair and plasticity |
| Microglia | Resident immune cells of the CNS | Survey tissue, respond to damage and pathogens, clear debris, regulate inflammation, and prune synapses |
| Ependymal cells and tanycytes | Cells lining ventricles and specialized CSF-facing interfaces | Line the ventricular system, help regulate the interface between brain tissue and cerebrospinal fluid, and in specialized regions participate in brain-body signaling |
| Radial glia / neural progenitors | Developmental scaffold and progenitor cells | Generate neurons and glia during development and guide migrating neurons into the developing cortex |
Two cautions go with the table. First, the living brain also contains blood-vessel cells, meningeal cells, immune-related cells at its borders, and connective-tissue elements. A living brain is not just a wiring diagram. Second, these categories are coarse. Modern single-cell methods can subdivide neurons and glia into dozens or hundreds of molecularly defined types. That is genuine progress, but a first-year student needs the major bins before the exquisite subdivisions. Otherwise the cell atlas becomes a phone book.
17.2 Neurons as excitable, polarized cells
A neuron is a cell specialized for receiving, integrating, and transmitting signals, and its anatomy reflects those three jobs. The soma, or cell body, contains the nucleus and much of the metabolic machinery. Dendrites are the receiving processes. They branch out from the soma and collect inputs from other neurons. Many dendrites are studded with small protrusions called dendritic spines, which are common sites of excitatory synapses. The axon is the output process. It begins at a specialized region called the axon initial segment, where action potentials are usually initiated, and it carries the neuron’s output to other cells. Near its end the axon branches into terminals with synaptic boutons, the small swellings where transmitter is released.
This arrangement gives the neuron functional polarity. The canonical flow is dendrites to soma to axon: input, integration, output. That rule is useful even though the biology is richer. Dendrites can perform local computations. Axons can give off collaterals that influence nearby circuits. Some synapses send signals backward from the postsynaptic cell to the presynaptic terminal. Electrical synapses can couple cells bidirectionally. But the polarized neuron remains the right starting image. It is the clean model we need before learning all the ways real neurons make the model more interesting.
The junction where one neuron’s axon communicates with another cell is a synapse, a word we owe to Charles Sherrington, who inferred the existence of such junctions from the logic of reflexes before anyone could see them clearly. The next chapter will explain how that synaptic influence actually works. For now, the essential point is anatomical: neurons have parts, the parts are arranged directionally, and that arrangement gives the cell a built-in logic.
17.3 The pyramidal neuron as the canonical example
The best first example is the pyramidal neuron of the cerebral cortex. It has a roughly triangular soma, a long apical dendrite that ascends toward the cortical surface, a branching apical tuft near the top, and several basal dendrites that spread out from the base of the soma. Its axon usually leaves from the basal side and may project to nearby cortical neurons, to other cortical areas, to subcortical targets, or even down toward the spinal cord, depending on the cortical region and layer.
This shape is not decorative. It is a functional arrangement. Basal dendrites sample one set of inputs, often local and feedforward. The apical dendrite and tuft sample another set, often including feedback, modulatory, or long-range inputs arriving in superficial cortical layers. That distinction should not be turned into a rigid rule, but it is the right intuition: a pyramidal neuron is built to compare and integrate signals arriving from different sources and at different distances from the soma.
Pyramidal neurons are also the major excitatory projection neurons of the cerebral cortex. They typically release glutamate and send long axons to other targets. Much of what students first learn about cortical signaling is really about pyramidal neurons and the inhibitory interneurons that control them. That is a reasonable starting bias, but it is a bias. The brain contains many neurons that do not look like pyramidal neurons, and the differences are not aesthetic. A difference in structure is usually a clue to a difference in function.
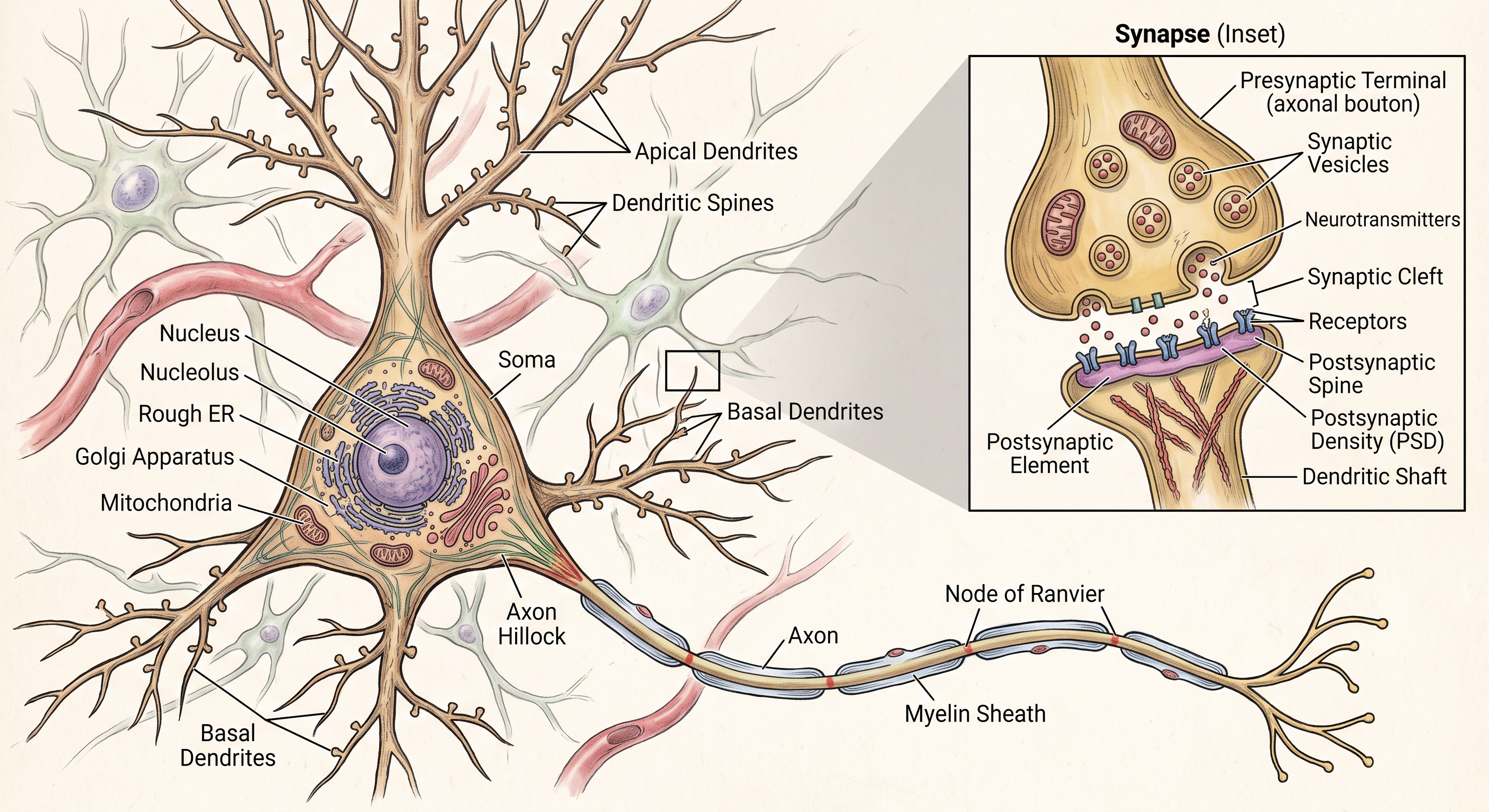
Synapse Inset (Top Right): This magnified view details a chemical synapse, where an action potential triggers neurotransmitter release. An axonal terminal (or ‘bouton’) from another neuron forms the presynaptic terminal. Within it are synaptic vesicles filled with neurotransmitters, along with supporting mitochondria. In response to a signal, these vesicles fuse with the presynaptic membrane and release their contents into the narrow, extracellular gap known as the synaptic cleft. The neurotransmitters then diffuse across the cleft and bind to specific receptors on the membrane of the target cell. In this example, the signal is received by a post-synaptic spine (a type of dendritic spine) attached to a dendritic shaft. The specialized area of the post-synaptic membrane, rich in receptors and signaling proteins, is identified as the postsynaptic density (PSD).
17.4 Not all neurons look like pyramidal neurons
Pyramidal neurons are a canonical example, not a universal template. Other brain regions have their own characteristic cells, and some of the most important neurons in the nervous system violate the simplified rule that “projection neurons excite and interneurons inhibit.”
| Neuron type | Main location | Typical message | Morphological signature | Functional implication |
|---|---|---|---|---|
| Pyramidal neuron | Cerebral cortex and hippocampus | Excitatory, usually glutamatergic | Triangular soma, apical dendrite, basal dendrites, many spines, long axon | Integrates inputs across cortical layers and projects to distant targets |
| Cortical interneuron | Local cortical circuits | Usually inhibitory, GABAergic | Diverse forms; often compact dendrites and local axonal arbors | Controls timing, gain, synchronization, and dendritic integration of nearby neurons |
| Medium spiny neuron | Striatum of the basal ganglia | Inhibitory, GABAergic | Medium-sized soma; dendrites densely covered with spines; projection axon | Integrates cortical, thalamic, and dopaminergic input for action selection |
| Purkinje cell | Cerebellar cortex | Inhibitory, GABAergic | Large soma; enormous, flattened dendritic tree | Integrates massive cerebellar input and provides the output of cerebellar cortex |
| Granule cell | Cerebellum, dentate gyrus, and other regions | Often excitatory, but region-dependent | Very small soma; compact dendrites; highly numerous in some regions | Allows dense recoding and expansion of input patterns |
Cortical interneurons deserve special warning because they are often described too casually as “small local inhibitory neurons.” Some are basket cells, whose axons wrap around the soma of pyramidal neurons. Some are chandelier cells, whose axon terminals target the axon initial segment, giving them extraordinary leverage over whether the target cell fires. Some, such as Martinotti cells, send axons toward distal apical dendrites and influence the inputs arriving there. Interneurons are specialized control elements, placed with precision. The composition of neurons within primate cerebral cortex is approximately 75-80% pyramidal neurons with interneurons making up the remainder. This represents a greater proportion of inhibitory interneurons than is found in rodent cortex where the proportion is closer to 85% pyramidal neurons.
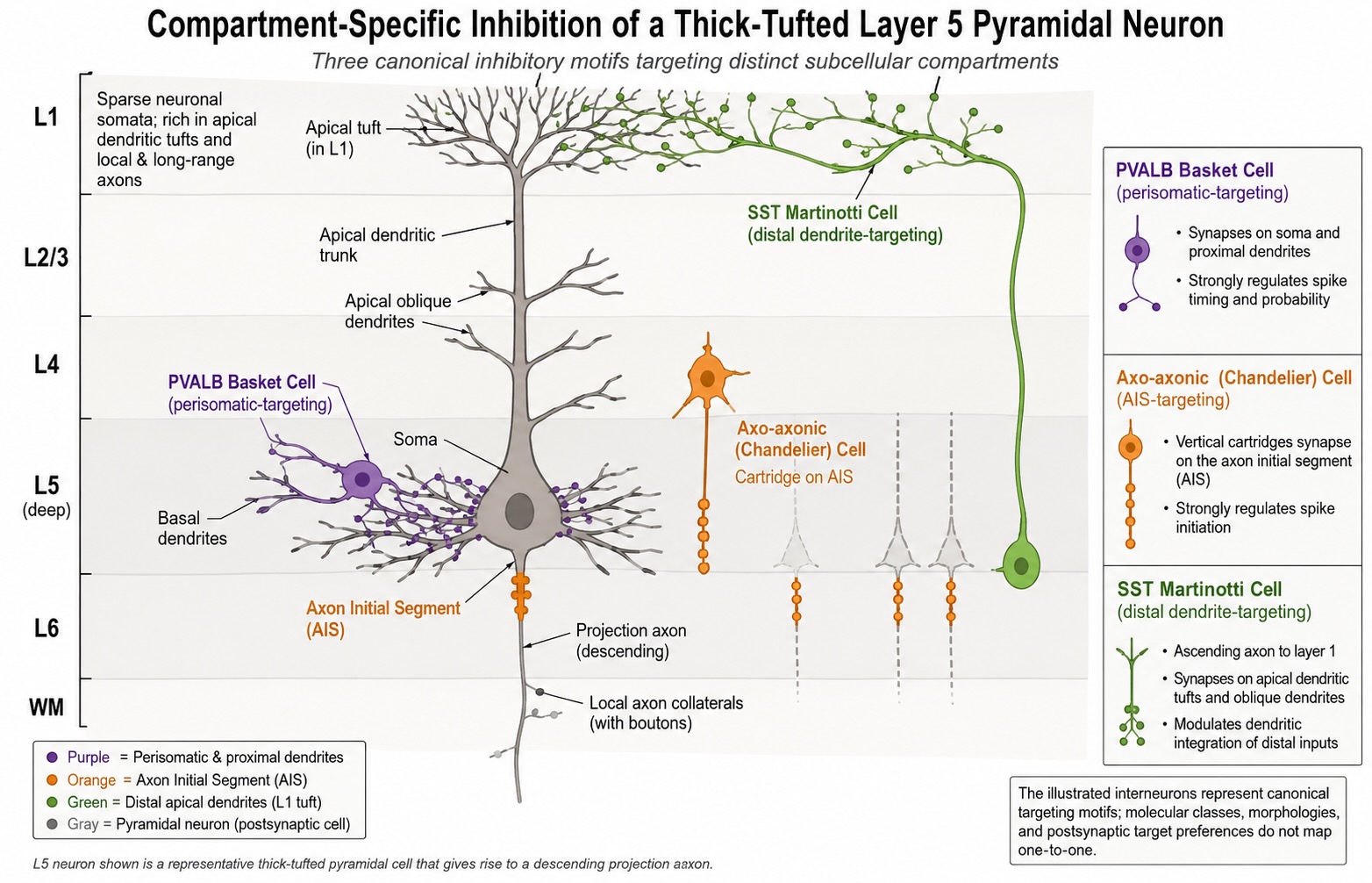
The same point applies across regions. Medium spiny neurons are the principal neurons of the striatum, but they are inhibitory. Purkinje cells are the principal output neurons of the cerebellar cortex, but they are inhibitory too. These cells are not exceptions that break the rules; they reveal that the rules were too simple. To classify a neuron well, we need several axes at once: what transmitter it releases, how far its axon travels, what shape it has, where it lives, what genes it expresses, and what circuit job it performs.
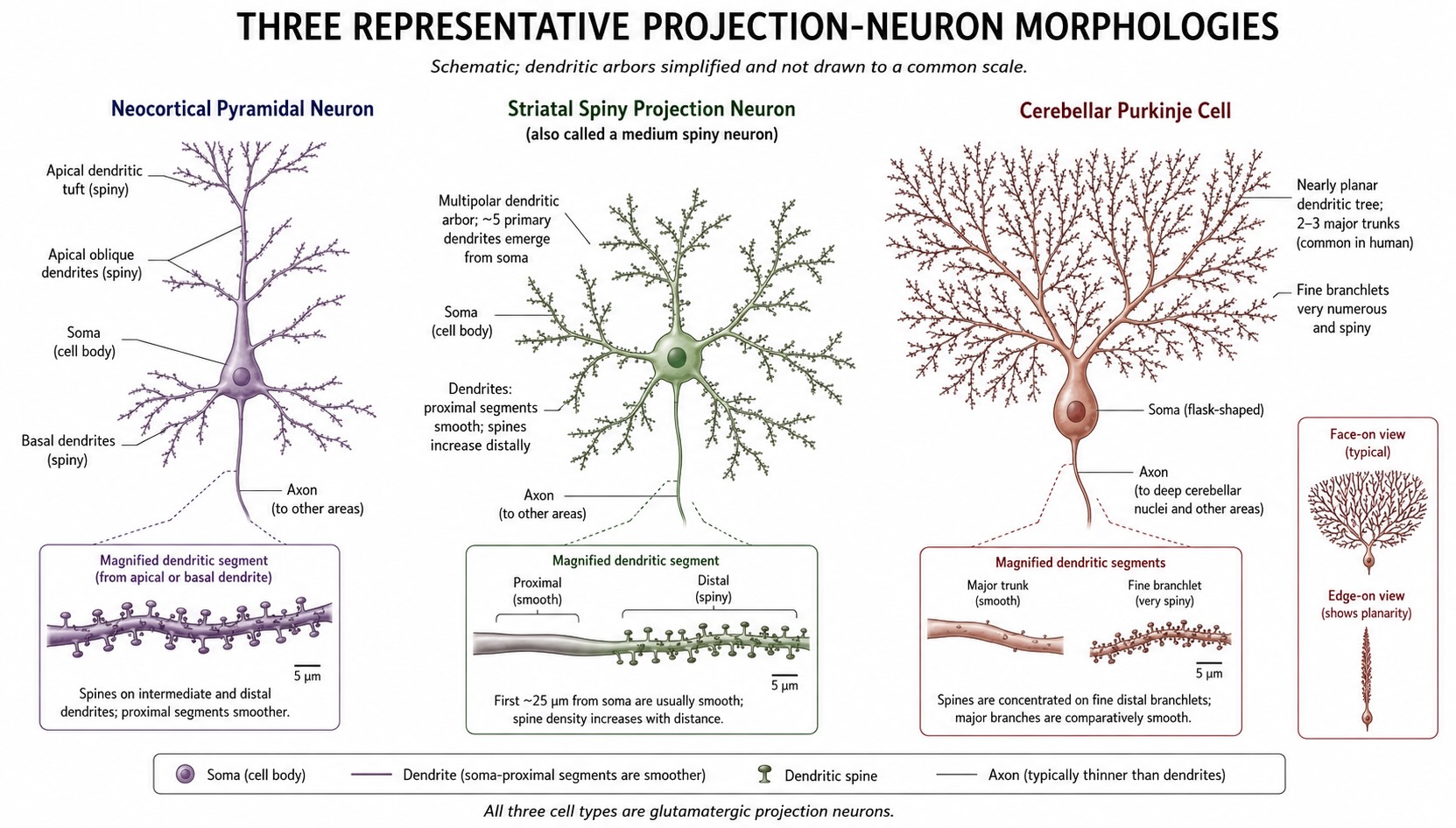
The idea that the brain is built from discrete neurons — separate cells, not a continuous fused network, or reticulum — was not obvious. In the late nineteenth century two great anatomists looked at the same kind of stained tissue and drew opposite conclusions. Camillo Golgi had invented the silver-impregnation stain that, for reasons still not fully understood, blackens a small random fraction of neurons in their entirety, revealing their full shape against a clear background. Golgi himself concluded that the nervous system was a continuous reticulum — a fused web in which the processes of different cells joined into one network. Santiago Ramón y Cajal, using Golgi’s own stain with extraordinary patience and artistry, came to the opposite view: that neurons are discrete cells, contacting one another closely but not fusing. The two shared the Nobel Prize in 1906, and Golgi famously used his Nobel lecture to attack Cajal’s position.
What I find incredible is that neither man could see the point of contact between two neurons. The gap between them is on the order of twenty nanometers, far below the resolution of any light microscope. Cajal was arguing from the appearance of discreteness — from processes that seemed to end freely, from the way a stained cell filled completely and stopped — and from developmental and degenerative evidence. He was inferring a cellular boundary he could not observe. The synaptic cleft would not be visualized until the electron microscopy of the 1950s, in work by Sanford Palay, George Palade, Eduardo De Robertis, and others. Cajal was right, but he was right in advance of the evidence that settled the question.
The doctrine and its laws: Cajal’s view became the neuron doctrine. In its classical form it holds that the neuron is the fundamental structural and functional unit of the nervous system; that neurons are discrete cells, not a syncytium; that each is organized into specialized parts — dendrites, soma, axon; and that information flows through the cell in one direction, from dendrites to soma to axon and onward to the next cell. This last claim, sometimes called the law of dynamic polarization, was Cajal’s great functional insight: it turned anatomy into circuitry. Once you accept that signals enter here and leave there, a tangle of stained cells becomes a wiring diagram, and you can start asking what the wiring does.
That is the doctrine’s real achievement. The doctrine made the nervous system analyzable. It licensed the whole subsequent enterprise of tracing pathways, identifying inputs and outputs, and modeling circuits as directed graphs of signal flow.
Exceptions have softened every claim: But each of the doctrine’s specific claims, taken as a strict law, has turned out to be an overstatement.
Discreteness is not absolute. Electrical synapses, formed by gap junctions, connect the cytoplasm of two neurons directly through channels that pass ions and small molecules. Current flows through them in both directions, and coupled cells can behave, electrically, almost as one. Golgi was wrong about the general architecture of the brain, but he was not wrong that neurons can be continuous with one another. Gap junctions are widespread — in the retina, in the inferior olive, among cortical interneurons — and they matter functionally, not as a curiosity.
Polarization is not strict. Information does not only run dendrite → soma → axon. Action potentials propagate backward into the dendritic tree, where they signal to the dendrites that the cell has fired — a mechanism central to synaptic plasticity, which we will meet again. Retrograde transmission carries signals backward across the chemical synapse itself: endocannabinoids and nitric oxide, released from the postsynaptic cell, act on the presynaptic terminal to change how much transmitter it releases. Some neurons release transmitter from their dendrites. Some axons receive synapses. The arrow in the textbook diagram is a statistical tendency, not a rule.
The neuron is not a single functional unit. Cajal’s doctrine treats the cell as the atom of computation: inputs sum at the soma, and the cell either fires or does not. In fact many dendrites are electrically active, with voltage-gated channels that support local spikes. A dendritic branch can perform its own nonlinear operation on the inputs it receives and pass on a result the soma never sees in raw form. The neuron looks less like a summing point and more like a tree of semi-independent compartments — a small network in its own right. And in the other direction, some cells (certain retinal amacrine cells, for instance) do not fire action potentials at all, and some processes are so short that the whole dendrite–soma–axon distinction becomes strained.
Nor is the neuron the only functional element in the tissue. Astrocytes respond to and modulate synaptic transmission; the classical doctrine treats glia as scaffolding, and they are not.
How to view the neuron doctrine today: There is a temptation to say that the neuron doctrine is “essentially correct, with exceptions.” That formula is too comfortable. If every one of a theory’s laws admits systematic exceptions, a better description is not that the doctrine is correct, but that it was a first approximation — and a spectacularly productive one.
The distinction matters. The neuron doctrine’s core structural claim — that the brain is built of cells, and that those cells are the units you should be counting — survives intact. What has not survived is the strict functional program that was projected onto it: the neuron as an indivisible unit of computation, with one input surface, one output, and one direction of flow. That picture was the right thing to believe in 1900. It gave neuroscience a tractable object and a century of results. It is not a description of how the machinery actually works.
17.5 Where cortical neurons come from
The first unit told the story of the brain in evolutionary and developmental terms together — evo-devo — and argued that the architecture we find is the architecture development builds. Nowhere is that clearer than in how the cortex is assembled, because two great classes of cortical neuron are born in different places and travel to their destinations by different routes. The classification we are about to make rests partly on this divergence, so it is worth seeing the developmental story before reducing neurons to labels.
The excitatory neurons of the cortex — the pyramidal cells we have just met — are born in the ventricular and subventricular zones lining the fluid-filled ventricles. From there they migrate radially: they climb more or less straight outward toward the surface, guided by the long fibers of radial glia, specialized progenitor-and-scaffold cells whose processes span the thickness of the developing cortical wall. The cortex is built inside-out. The first neurons to arrive settle in the deepest layers, and each later wave migrates past the cells already in place to settle more superficially, so that the youngest neurons end up nearest the surface. This sequence — deep layers first, superficial layers last, each generation climbing past its elders — is the developmental signature of the cortex’s layered structure.
The inhibitory neurons take an entirely different path. Cortical interneurons are not born in the cortex at all. Many originate in the ganglionic eminences, transient structures in the ventral part of the developing forebrain, and they reach the cortex by migrating tangentially — traveling long distances sideways across the developing brain before turning to integrate into local cortical circuits. Two classes of neuron that sit together in adult cortex therefore have very different biographies. The excitatory pyramidal neuron is born locally and climbs outward. The inhibitory interneuron is born elsewhere and immigrates.
That difference matters. It helps explain why the cortical excitatory population is relatively uniform in broad outline — pyramidal cells, layered by birthdate and projection pattern — while inhibitory interneurons are extraordinarily diverse. It also gives us a developmental handle on disease. If radial migration is disrupted, neurons can end up stalled near the ventricles, producing periventricular nodular heterotopia, or the cortex can fail to form its normal folds and layers, producing forms of lissencephaly. If tangential migration or interneuron maturation is disrupted, the balance of excitation and inhibition can be altered, with consequences that may include epilepsy and neurodevelopmental disorders. The adult circuit carries the history of how its cells got there.
This is also where radial glia deserve their first serious mention. They are called glia, but in development they are not merely support cells. They generate both neurons and glia, provide scaffolds for migration, and then, in many regions, transform into astrocytic lineages. Development keeps making the same point: the categories are real, but the boundaries are not as tidy as textbook tables make them look.
17.6 Classifying neurons: message, reach, shape, and molecules
Once the cells have arrived, how should we classify them? There is no single correct answer because neurons differ along several useful axes. The trick is not to memorize one taxonomy, but to know which axis is being used.
The first axis is the message. A neuron may be excitatory, usually releasing glutamate, or inhibitory, usually releasing GABA. Excitation pushes the target cell closer to firing. Inhibition pushes it farther from firing or shunts incoming current so that excitation has less effect. This is the simplest circuit vocabulary: excite or inhibit.
The second axis is reach. Projection neurons have axons that carry information to distant targets. Interneurons act locally within a circuit. This axis is anatomical, not chemical. A neuron can be long-range and excitatory, long-range and inhibitory, local and inhibitory, or, in some circuits, local and excitatory. In cerebral cortex the common pattern is that pyramidal projection neurons are excitatory and local interneurons are inhibitory, but the brain is not obligated to keep that pattern everywhere. The basal ganglia and cerebellum immediately teach otherwise.
The third axis is shape. Pyramidal cells, medium spiny neurons, Purkinje cells, granule cells, basket cells, and chandelier cells are named partly because they look different. Morphology is not superficial. It tells you where the cell receives input, where it sends output, how many partners it may contact, and what kind of computation it is positioned to perform. A Purkinje cell’s immense flattened dendritic tree is the physical solution to a particular cerebellar problem.
The fourth axis is molecular identity. This is where the modern taxonomy has become especially powerful. Many inhibitory interneurons can be grouped by the molecules they express. Three important cortical classes are parvalbumin-positive (PV), somatostatin-positive (SST), and VIP-positive interneurons.
PV interneurons include fast-spiking basket cells and chandelier cells. They are well positioned to control the timing and probability of pyramidal-cell firing. SST interneurons often target dendrites, including distal dendrites, and can regulate what inputs are allowed to influence a pyramidal cell. VIP interneurons often inhibit other inhibitory neurons, producing disinhibition: they can increase activity in a target population by turning down the local cells that suppress it. That sounds like a trick, but it is one of the brain’s common strategies. Inhibiting an inhibitor is a way of opening a gate.
This molecular way of thinking should feel familiar from the hypothalamus. The arcuate nucleus contains intermingled populations such as AgRP neurons and POMC neurons that sit side by side yet pull feeding behavior in opposite directions. What separates them is not an obvious gross shape but a molecular signature and a circuit role. The same logic applies in cortex, basal ganglia, brainstem, and many other regions. When shape and location are not enough, molecular identity can reveal functional differences that anatomy alone would miss.
Here is a caution. Single-cell transcriptomics has produced cell-type taxonomies of extraordinary fineness — dozens or hundreds of putative neuronal and glial types defined by gene-expression profiles. This is one of the genuine revolutions of modern neuroscience. It is also not a finished catalog handed down from the mountain. Some “types” may be discrete biological classes; others may be points sampled along continuous gradients of variation. The three-marker interneuron scheme is a robust coarse-graining. The finer structure beneath it is real, powerful, and still being argued about.
17.7 From single neurons to local circuits
The next chapter will go into the details of how neurons actually excite and inhibit their targets: membrane potentials, ion gradients, action potentials, synaptic release, receptors, and the ways astrocytes modulate the synaptic environment. For now, we can stipulate the basic output rule. A neuron influences its target by excitation, inhibition, or slower modulation of those effects.
That stipulation lets us take a step upward. Neurons are not isolated devices. They exist in circuits. Once one neuron can excite or inhibit another neuron, we can begin to build small circuit patterns. These patterns are called motifs: recurring arrangements that appear in many neural systems. A motif is not a whole brain function. It is a reusable piece of circuit grammar.
One of the earliest formal attempts to think this way came from Warren McCulloch and Walter Pitts. In 1943, they published A Logical Calculus of the Ideas Immanent in Nervous Activity, a paper that treated neurons as simplified logical units. A McCulloch-Pitts neuron received inputs, summed them, and produced an all-or-none output if the input exceeded a threshold. By connecting such units into networks, McCulloch and Pitts showed how neural circuits could implement logical operations. Their neurons could be arranged to perform operations resembling AND, OR, and NOT gates, which are the fundamental Boolean operations of computation.
Their model was biologically crude. Real neurons have dendrites, synapses of different strengths, delays, nonlinear membranes, spontaneous activity, modulators, plasticity, noise, and metabolic constraints. Real neurons are not little Boolean beads on a wire. Still, the abstraction was powerful because it made a crucial idea explicit: functions can be built from networks of simple units whose interactions produce something not present in any single unit. That idea runs through computational neuroscience, artificial neural networks, and much of modern systems neuroscience.
For this course, the point is simpler and more biological. If neurons can excite and inhibit, then we can build circuit motifs from those two operations.
In the diagrams below, → means excitation and ┤ means inhibition.
| Motif | Simple diagram | What it does |
|---|---|---|
| Feedforward excitation | A → B → C | Carries a signal forward from one processing stage to the next |
| Feedforward inhibition | A → B and A → I ┤ B | Lets an input excite a target while also recruiting inhibition that limits the time window of that excitation |
| Feedback / recurrent inhibition | B → I ┤ B | Lets an active neuron or population recruit inhibition that stabilizes its own activity |
| Lateral inhibition | A → B and A → I ┤ neighboring B | Sharpens contrast by suppressing neighboring pathways |
| Recurrent excitation | A → B → A | Amplifies activity and can support persistent states, but requires inhibition to prevent runaway excitation |
| Disinhibition | A ┤ I ┤ B | Inhibits an inhibitor, thereby releasing a target from inhibition; a common gating strategy |
| Convergence | A, B, C → D | Combines multiple inputs onto one neuron or population |
| Divergence | A → B, C, D | Broadcasts one signal to multiple targets |
These motifs are not tiny machines with only one use. They are patterns that can be reused in different anatomical settings. Feedforward inhibition will appear in sensory pathways where timing matters. Lateral inhibition will appear when the nervous system sharpens spatial contrast, as in the retina and somatosensory system. Disinhibition will appear in cortical microcircuits and basal ganglia loops. Recurrent excitation will appear whenever we ask how activity can persist long enough to guide behavior, and recurrent inhibition will appear whenever we ask why that activity does not simply explode.
This is why a cellular chapter belongs before the sensory systems. The retina, cochlea, thalamus, cortex, and spinal cord are not just chains of anatomical relays. They are circuits built from excitation, inhibition, convergence, divergence, feedback, and modulation. Learning the motifs now gives us a vocabulary for recognizing them later, when they are embedded in real sensory anatomy.
17.8 Glia: not glue, and not optional
We have spent most of the chapter on neurons because neurons provide the fast excitable signaling that the next chapters will analyze. But roughly half the cells in the brain are not neurons, and the old dismissal of them as glia — glue — has not survived. Glia do not usually fire action potentials, but not all important signaling in the brain looks like a spike train.
It is useful to separate the major glial classes before focusing on astrocytes.
17.8.1 Oligodendrocytes, myelin, and OPCs
Oligodendrocytes are the myelinating cells of the central nervous system. They wrap segments of axons in myelin, a fatty insulating sheath that makes action potentials travel faster and more reliably. The next chapter will explain why myelin changes conduction so dramatically. For now, the anatomical fact is enough: an oligodendrocyte can myelinate segments of multiple axons, and the myelin sheath is interrupted by gaps called nodes of Ranvier, where action potentials are regenerated.
The peripheral nervous system uses a related but different glial cell, the Schwann cell, to myelinate peripheral axons. Schwann cells are not brain glia, but both cell types solve the same electrical problem by wrapping axons in insulation.
A related class, oligodendrocyte precursor cells, often called OPCs or NG2 glia, persists in the adult brain. The name makes them sound like cells waiting passively to become oligodendrocytes, but that is too narrow. They can generate new oligodendrocytes, respond to injury, and participate in activity-dependent changes in myelination. This matters because myelin is not just developmental wiring put in place once and then forgotten. The speed and reliability of signaling can be altered by changes in myelination, and OPCs are part of that plastic machinery.
17.8.2 Microglia: immune cells that sculpt circuits
Microglia are the resident immune cells of the central nervous system. They are different from astrocytes, oligodendrocytes, and ependymal cells not only in function but in origin. The macroglia are neuroectodermal; microglia come from an immune lineage and enter the developing nervous system early. They are brain residents, but they are not born from the same developmental family as the other glia.
Microglia survey neural tissue with fine, motile processes. They respond to injury and infection, clear debris, release inflammatory signals when needed, and help maintain tissue health. That would already make them important. But they also help sculpt circuits. During development, the brain produces more synaptic connections than it will keep. Some synapses are stabilized; others are eliminated. Microglia participate in removing some of those connections. The immune cell of the brain is therefore also a circuit-shaping cell.
During development, the brain overproduces synaptic connections. Some are stabilized; others are removed. One of the important discoveries of modern neuroscience is that part of this removal is performed by microglia, the resident immune cells of the central nervous system.
Microglia are motile and phagocytic. In the developing brain they extend and retract fine processes, sampling the local environment. When synapses are weak, inappropriate, damaged, or molecularly marked for elimination, microglia can engulf synaptic material. This process is often called synaptic pruning.
The immune system provides part of the tagging machinery. Molecules of the complement system, including C1q and C3, can mark synaptic elements for removal. Microglia express complement receptors and can recognize these tagged structures. A molecular system associated with immune defense is therefore reused by the developing brain to refine circuits.
This does not mean that microglia randomly wander around eating synapses. Pruning is regulated by development, neural activity, molecular tags, and local context. Nor is microglial function limited to childhood. In the adult brain, microglia continue to survey tissue and can participate in remodeling, inflammation, disease, and possibly some forms of forgetting. But the developmental lesson is the cleanest: building a brain requires demolition as well as construction.
17.8.3 Ependymal cells, tanycytes, and radial glia
Ependymal cells line the ventricles, the fluid-filled spaces inside the brain. Their cilia help move cerebrospinal fluid. They are not usually the stars of an introductory neuroscience chapter, but they matter because the brain is bathed by fluid and separated from that fluid by living cellular interfaces, not inert plastic wrap.
Specialized ventricular cells called tanycytes, especially near the hypothalamus, help regulate interfaces between cerebrospinal fluid, brain tissue, and body-state signals. This connects the cellular chapter back to the hypothalamus and circumventricular organs. The nervous system does not only receive information through eyes and ears. It also samples the state of the body, and some of that sampling occurs at interfaces between blood, cerebrospinal fluid, and neural tissue.
Radial glia are most important in development. They are progenitor cells and scaffolds: they generate neurons and glia, and their long fibers guide migrating neurons into position. Calling them “glia” can therefore be misleading. In development, radial glia are part of the biological machinery that builds the nervous system.
17.8.4 Astrocytes: the operating environment of synapses
Astrocytes deserve extended treatment because they change how we should imagine the synapse. Of all the glia, they are the ones most likely to make the standard cartoon of neural signaling look inadequate. Oligodendrocytes change the speed of axons. Microglia maintain and remodel circuits. Astrocytes change the local operating conditions under which synapses work.
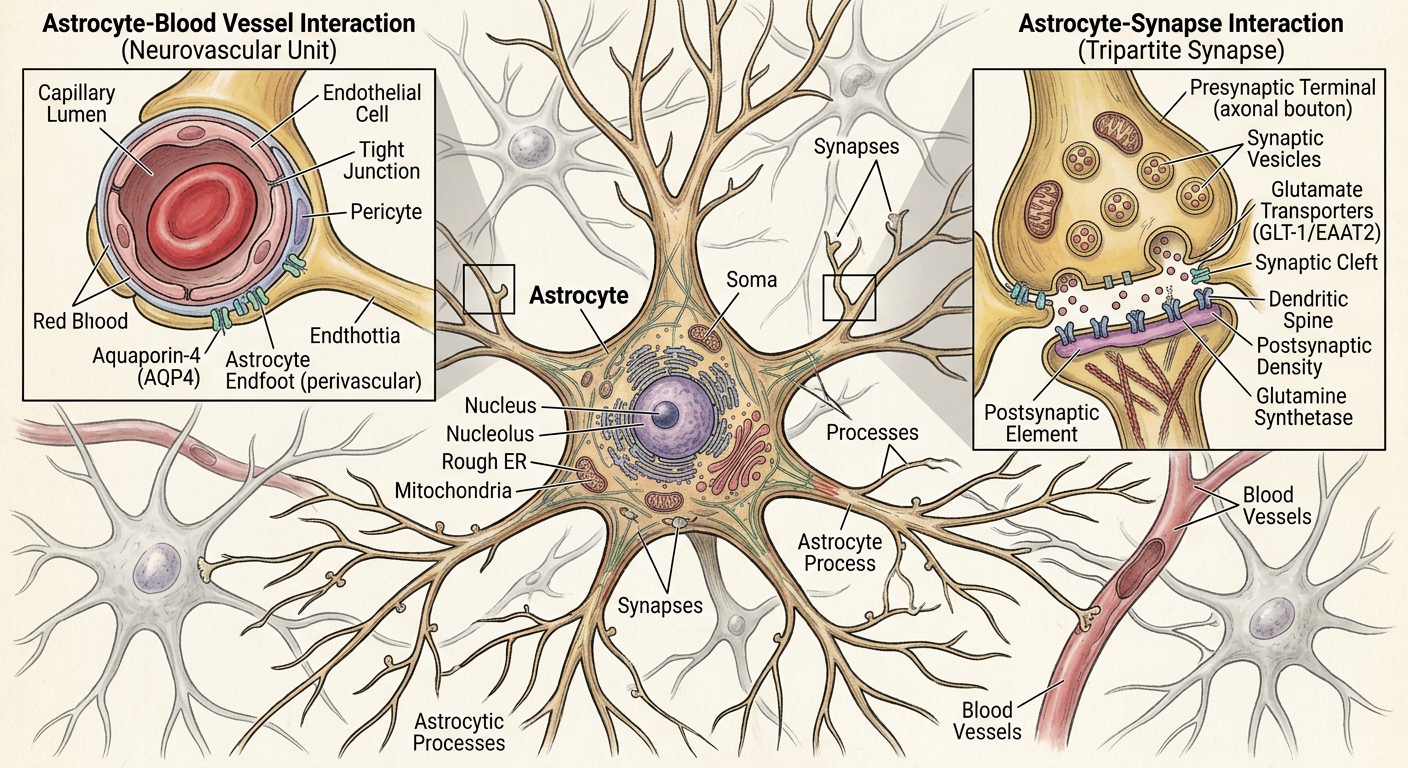
The classical synapse has two parties: the presynaptic terminal that releases transmitter and the postsynaptic membrane that receives it. The modern picture adds a third. Astrocyte processes wrap closely around many synapses, sensing and shaping what happens there, and the assembly of presynaptic terminal, postsynaptic membrane, and astrocytic process is often called the tripartite synapse. The astrocyte is not a passive bystander at the junction. It does work that bears directly on signaling.
The most basic work is housekeeping that turns out not to be basic at all. When a glutamatergic synapse fires, glutamate floods the cleft, and it must be cleared promptly — both to sharpen the signal and because glutamate left lingering in the extracellular space is excitotoxic, capable of overexciting and killing neurons. Astrocytes take up excess glutamate, convert it to glutamine, and ship the glutamine back to the presynaptic neuron to be recycled into fresh transmitter. They also buffer the potassium that accumulates outside neurons during bouts of firing, helping keep the extracellular environment stable. These are not glamorous functions, but without them fast signaling would poison its own surroundings.
Astrocytes also sit at a metabolic crossroads. They extend specialized endfeet that wrap around the brain’s blood vessels, and through them astrocytes help couple local neural activity to local blood flow — the phenomenon called neurovascular coupling. This coupling is much of what brain-imaging methods like fMRI actually measure: not neural activity directly, but blood-flow and oxygenation changes that track it. Astrocytes are therefore part of the bridge between synaptic activity and the energy supply that supports it.
A single astrocyte can contact a very large number of synapses, which places it in an ideal position to sense local demand and help regulate local conditions. The exact numbers vary by species, brain region, and method, but the conceptual point is secure: astrocytes are positioned at the intersection of synapses, extracellular chemistry, metabolism, and blood vessels. The cell once dismissed as glue turns out to be a recycler, a buffer, a metabolic partner, a vascular intermediary, and possibly a participant in signaling.
17.9 Gliotransmission and the calcium-wave debate
Astrocytes are coupled to one another through gap junctions, forming networks through which signals can spread. Their arrangement recalls Golgi’s concept of a reticulum, but for astrocytes and not neurons. The signal in question is also not the same as the action potential of a neuron. Astrocytes often signal through changes in intracellular calcium, which can occur locally in fine processes or more broadly across the cell. This raised a genuinely interesting possibility: perhaps astrocytes do not merely support neuronal signaling but participate in it, releasing their own signaling molecules back onto neurons.
That possibility is called gliotransmission, and it has had a turbulent history. It is a good example of how neuroscience should work: an exciting idea appears, expands too broadly, gets criticized, and then becomes more precise.
The basic proposed mechanism runs like this. Glutamate released at a synapse can activate receptors on nearby astrocytic processes, raising calcium inside the astrocyte. This calcium increase can remain local, spread through astrocyte processes, or propagate through networks of astrocytes coupled by gap junctions. Under some conditions, the astrocyte may then release signaling molecules — sometimes called gliotransmitters — that influence nearby neurons.
Candidate gliotransmitters include glutamate, ATP, GABA in some contexts, and D-serine. D-serine is especially interesting because it is a required co-agonist at the NMDA receptor, the coincidence-detecting receptor central to many forms of synaptic plasticity. An astrocyte that regulates local D-serine availability could therefore influence whether nearby synapses are capable of strengthening.
The controversy is about how general, how fast, and how behaviorally important this mechanism is in the intact adult brain. Some early demonstrations used immature tissue, cell culture, or stimulation conditions that may not reflect normal adult physiology. Some receptors central to the original models are much more prominent in developing tissue than in adult astrocytes. Astrocytic calcium signals can also be slower than neuronal electrical signals, which makes them unlikely to explain every rapid synaptic effect once attributed to them.
The field has not simply abandoned the idea. It has narrowed it. Astrocytes have compartments, and small local calcium events in fine processes may matter differently from large calcium waves in the soma. Astrocytes can influence synapses and plasticity, but not every strong claim of fast glial control has survived. That is not a defeat, but rather the normal sharpening of a scientific idea.
The important conceptual point is not that astrocytes are secretly neurons. They are not. They usually do not send millisecond action potentials down axons. Their signaling is often slower, more local, and more biochemical. The point is that neurons do not signal in a vacuum. The synapse is embedded in tissue, and astrocytes help set the local operating conditions of that tissue.
17.10 Looking ahead
We now have the cast. Neurons are excitable signaling cells with polarized morphology. Pyramidal neurons, interneurons, medium spiny neurons, Purkinje cells, and granule cells differ in shape because they differ in job. Cortical excitatory and inhibitory neurons have different developmental origins, and that developmental history helps explain adult circuit organization. Neurons can be classified by message, reach, shape, molecular identity, and circuit role. Glia are not glue. They regulate, insulate, defend, prune, guide, recycle, and metabolically support neural tissue.
We have also taken the first step from cells to circuits. Once neurons can excite and inhibit one another, motifs such as feedforward inhibition, lateral inhibition, recurrent inhibition, convergence, divergence, and disinhibition become possible. These motifs will reappear in the sensory systems, where they become concrete anatomical circuits rather than abstract diagrams.
What we have not yet done is explain how excitation and inhibition actually happen. The next chapter takes up that fast electrical layer in earnest: membranes, ion gradients, channels, action potentials, synapses, and receptors. After that, we will widen the frame again to neurotransmitters, neuromodulators, volume signaling, and the chemical anatomy of the brain. Finally, we will turn to how neurons learn from experience.