30 Action: Controlling a Body in a World
Overview: The Control System in Action
30.1 From sensing to action
The argument begun in Chapter 1 has now assembled most of a control loop. The hypothalamus and related systems introduced in Chapter 6 monitor the regulated body, anticipate needs, and alter the organism’s physiological state. The sensory chapters then turned outward, while Chapter 22 showed how estimates of the body’s own configuration remain inseparable from movement. Memory preserves information about what happened before, and learning changes what the same situation will mean next time. None of those operations, by itself, changes the animal’s relation to the world.
Action closes the loop.
A thirsty animal turns toward water, walks to it, lowers the head, opens the mouth, and swallows. A cold animal moves into the sun or curls its body to reduce heat loss. A hand reaches toward a cup, shapes itself around the handle, lifts against an uncertain load, and keeps the liquid level while the arm moves. In each case, neural activity becomes consequential by changing a body, and the changed body alters the sensory evidence arriving next.
The claim needs one boundary. Not every regulatory response ends in skeletal muscle. Earlier chapters described autonomic and endocrine effectors: blood vessels constrict, brown fat produces heat, kidneys retain water, and hormones alter tissues throughout the body. This unit concentrates on somatic action—posture, locomotion, orienting, reaching, grasping, speaking, and other behavior produced through skeletal muscle. Somatic action is not the only output of the brain, but it is the principal means by which an animal changes where it is, what it contacts, and what happens next in the external world.
The word action also deserves more care than the ordinary synonym movement. A movement is a physical change in the body. An action is movement—or the controlled maintenance or withholding of movement—organized with respect to an outcome. Tremor is movement without being a chosen action. Holding a tray level is an action even when the hands remain nearly still. Pressing against an immovable surface produces force without displacement. Stopping a prepared response can be as important as releasing it. The outcome need not be represented as a conscious intention: a withdrawal reflex or postural response can be organized around protecting tissue or preserving balance without deliberation. The distinction will matter throughout this unit because the nervous system does not merely generate motion. It stabilizes variables that matter to the animal while allowing many physical details to vary.
This overview therefore develops a deliberately broad claim:
Action is the hierarchical, closed-loop control of a biomechanical body in a changing world.
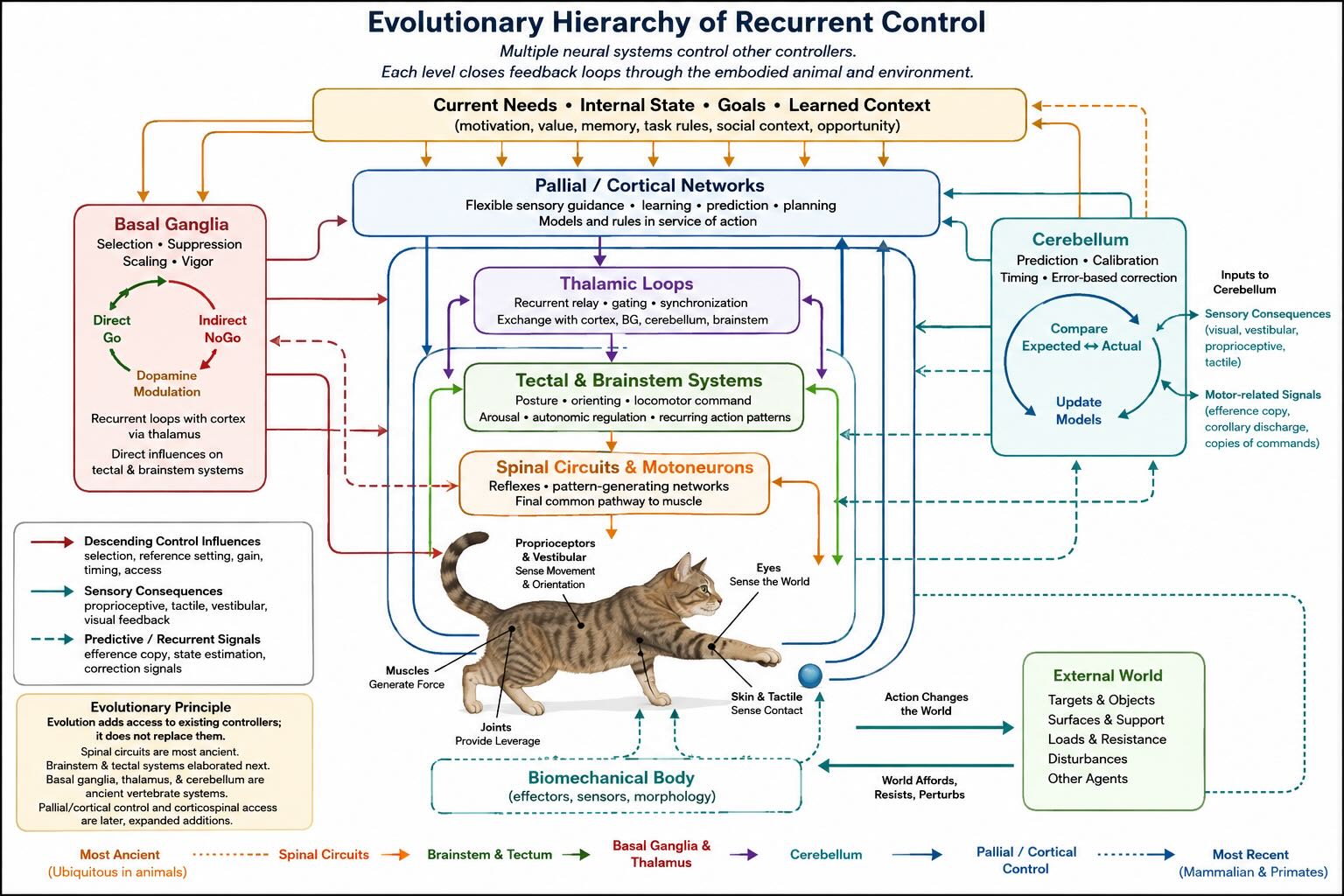
The hierarchy is real. Spinal and brainstem circuits can organize movement locally; forebrain systems select, redirect, suppress, and combine their outputs; cerebellar circuits calibrate predictions and corrections; and the expanding cerebral cortex adds increasingly flexible sensory guidance and learned control. But hierarchy does not mean a small executive issuing a complete muscle-by-muscle script. Each level is itself a controller. Later-evolving systems govern older controllers partly by changing their goals, gains, timing, and conditions of release. Sensory consequences then return through several routes and alter activity at every level.
Evolution built control above control without discarding what already worked [@grillnerelmanira2020; @grillner2021evolution].
30.2 Action is not the same as movement
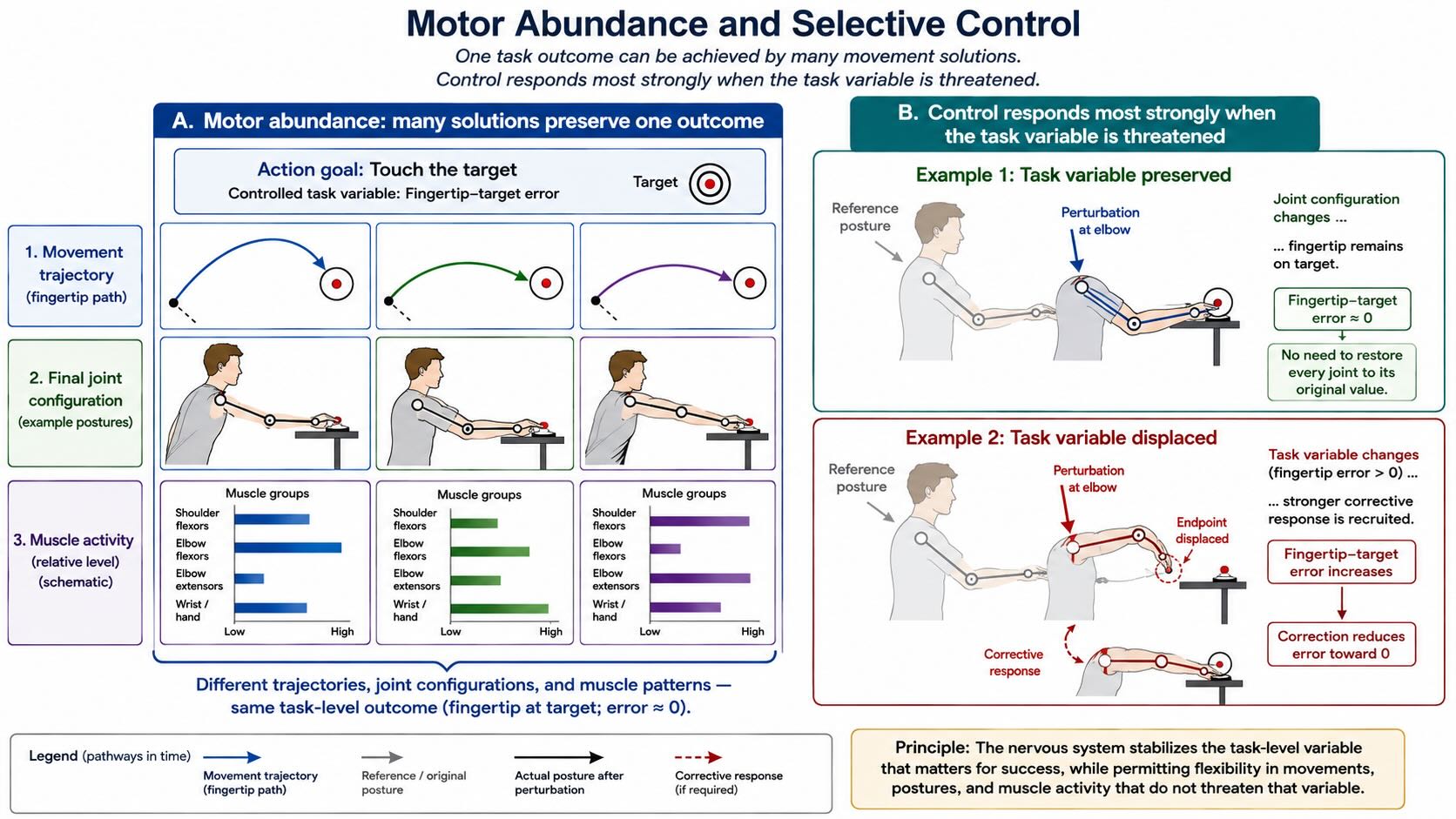
Touch the tip of your index finger to a point on the table. The task appears simple because the outcome is simple: fingertip at target. The movement that accomplishes it is not. The shoulder can rotate through several combinations, the elbow can flex by different amounts, the wrist can compensate, the trunk can lean, and the fingers can change configuration. A person can reach the same point from another initial posture, while holding an object, after fatigue, or with one joint partly constrained. No single pattern of joint angles or muscle activity is the reach.
This is the problem Nikolai Bernstein described as the degrees-of-freedom problem. A body contains many more controllable elements than are uniquely specified by most tasks. The nervous system could treat that abundance as a nuisance and attempt to freeze every unnecessary degree of freedom. Beginners sometimes do exactly that: a novice skier or pianist stiffens joints that an expert allows to move. Skilled control more often uses the abundance. Different combinations of joints and muscles can preserve the same important outcome, absorb perturbations, distribute fatigue, and adapt to injury [@bernstein1967coordination; @latash2012abundance].
The same principle appears at several levels. Several muscles can contribute to the same joint torque. Several joints can place the hand at the same location. Several trajectories can reach the same object. The right hand, left hand, foot, voice, or a tool can sometimes achieve the same environmental result. This capacity is called motor equivalence. It explains how a familiar action can survive changes in posture and effector without requiring the brain to retrieve one immutable motor file [@latash2012abundance].
For this reason, motor abundance is often a better term than motor redundancy. Redundancy sounds like useless duplication. Abundance supplies options. A controller can permit variation along dimensions that do not threaten the task while resisting variation along dimensions that do. When carrying a tray, for example, the absolute heights of the two hands can vary together without spilling anything; a difference in their heights tilts the tray and must be corrected. During a reach, elbow and shoulder angles can vary substantially if their covariation preserves the fingertip path. The nervous system does not need to correct every deviation equally. It intervenes most strongly where the deviation matters.
This minimum-intervention principle is one of the central insights of optimal-feedback accounts of motor control. It should not be turned into a rigid law: people sometimes respond to changes that an experimenter has defined as task irrelevant, and biological controllers carry habits, energetic costs, safety margins, and uncertainty that the laboratory task may not capture. The durable point is more modest. Good control is selective. It stabilizes outcomes rather than forcing every component of the body through one exact trajectory [@latash2002variability; @todorovjordan2002].
The distinction between action and movement also dissolves an apparent opposition between control and variability. Variability is not always noise. Some variability reveals failure, especially when it changes the task outcome. Other variability reveals flexibility: the system is exploiting multiple solutions while protecting what matters. An expert does not reproduce the same microscopic movement every time. The expert reproduces the result under changing conditions.
30.3 Movement is older than nervous systems
A nervous system did not invent the problem of action. Living cells already had to move toward favorable conditions, away from damaging ones, and around obstacles.
A bacterium swimming through a chemical gradient can bias its motion toward nutrients. It does not calculate a map. It compares recent conditions with slightly earlier ones. When conditions improve, it prolongs a relatively straight run; when they worsen, it tumbles and reorients. The result is a biased random walk that carries the cell, on average, toward what sustains it. A paramecium coordinates thousands of cilia, reverses their beat after contact, and changes direction. These are not conscious intentions, but they are organized actions in the biological sense: changes in motion coupled to conditions relevant to survival [@berg1972chemotaxis].
Nervous systems made this coupling faster, more extensive, and more flexible. They linked distant sensors to distant effectors, coordinated many contractile elements, and allowed present signals to be modified by internal state and prior experience. They did not replace the older principle. Sensors still alter effectors through physical circuits, and the consequences of action still change what the sensors report next.
Evolution is conservative in another sense. Once a circuit solves a recurring control problem, later evolution often modifies, duplicates, and places the older machinery under new forms of control rather than rebuilding the entire solution at a higher level. Vertebrate breathing did not become a sequence consciously composed in cortex. Skilled hand use did not require the spinal cord to stop processing stretch, load, and cutaneous input. A pianist’s cortex gains unusually refined access to the hand, but every note still depends on spinal interneurons, motoneurons, muscle mechanics, and sensory feedback that long predate pianos and primates.
This is where hierarchy enters the evolutionary story. The mistake is not to say that motor control is hierarchical. The mistake is to imagine that the hierarchy contains one true controller at its summit and passive machinery below. A spinal circuit can control muscle length while a brainstem circuit changes the operating conditions of that spinal circuit. A basal-ganglia loop can help release one brainstem action system while maintaining inhibition over competitors. Cortex can configure the selected action for a novel object and a social rule. Each higher level controls lower controllers rather than replacing control with command.
Comparative evidence indicates that much of this architecture was already present near the base of vertebrate evolution: spinal and brainstem pattern-generating systems, a tectum coupling mapped sensory events to orienting, basal-ganglia circuits involved in selection, developmental and circuit precursors of the cerebellum, and a pallium receiving and transforming sensory information. Mammalian evolution greatly expanded some components, particularly pallial and cortical systems and their access to distal musculature, but the expansion was a reweighting and elaboration of an inherited hierarchy, not a fresh motor system built above an inert cord [@grillner2021evolution; @ocana2015pallium; @sugahara2021cerebellum].
30.4 Pattern generators and local spinal control
One way to see local control clearly is to study a movement that repeats or unfolds as a structured sequence. Walking, swimming, breathing, chewing, scratching, and swallowing all require ordered activity across several motor populations. Sensory feedback can help pace and shape the sequence, but the movement cannot depend on each completed contraction serving as the sole instruction for the next. A useful pattern must be organized centrally.
A central pattern generator, or CPG, is a neural network capable, under suitable activation, of generating an ordered motor pattern when neither descending input nor sensory feedback supplies that pattern moment by moment. The definition is operational. It does not mean the network receives no input at all. A CPG may require tonic excitation, neuromodulators, or a trigger. It does not mean that the intact behavior normally runs without feedback. It does not require one continuously oscillating clock. The defining result is that the temporal organization can be generated within the network rather than copied from a rhythmic signal arriving from elsewhere [@grillner2003motor; @grillnerelmanira2020].
The classic starting model is the half-center oscillator proposed by Thomas Graham Brown. Two populations associated schematically with flexion and extension inhibit one another. Activity in one suppresses the other; adaptation, synaptic fatigue, post-inhibitory rebound, or other membrane and circuit properties eventually permit the suppressed side to take over. Alternation follows. The model is powerful because reciprocal inhibition is a recurring motif and because it shows how a pattern can emerge from relations among neurons rather than from a conductor [@brown1911progression].
The half-center is not the final anatomy of vertebrate locomotion. Real spinal networks contain interacting modules for rhythm generation, pattern formation, left–right coordination, flexor–extensor coordination, speed, and the distribution of activity across particular motor pools. Different species and behaviors implement these functions differently. The half-center should therefore be treated as a mechanism that makes a principle visible, not as a wiring diagram to be copied into every spinal segment [@grillnerelmanira2020].
Mutual inhibition by itself does not guarantee alternation. If two perfectly stable populations only inhibit one another, whichever gains an initial advantage might remain active indefinitely. A rhythm requires some process that weakens the active side or strengthens the suppressed side over time.
Several neuronal properties can provide that process. In spike-frequency adaptation, continued firing recruits currents that progressively reduce excitability. In post-inhibitory rebound, a neuron becomes especially likely to fire when prolonged inhibition is released. Persistent inward currents can sustain activity once it begins, while slower opposing currents eventually terminate it. Some neurons show intrinsic bursting, and synapses can facilitate or depress across repeated activity. Network rhythms can arise from combinations of these properties rather than from one dedicated pacemaker cell.
The resulting circuit is not executing a symbolic instruction that says “alternate.” Alternation is the trajectory produced by membrane properties, synaptic signs, connection strengths, and incoming drive. Change a neuromodulator and the same network may run faster, slower, or in a different mode. Change sensory input and a phase transition can be advanced or delayed. The architecture generates a family of patterns whose exact expression depends on state.
The strongest evidence for a CPG comes from preparations in which movement-generated feedback has been removed while patterned motor output remains. In fictive locomotion, an animal may be immobilized while rhythmic bursts are recorded from motor nerves. Because the limbs are not cycling, each flexion cannot be mechanically triggering the next extension. The pattern must be organized centrally. Comparable logic has revealed pattern-generating networks for swimming, respiration, scratching, and defensive acts such as vomiting [@grillner2003motor; @grillnerelmanira2020].
Spinalized mammals provide a different kind of evidence. After complete transection, cats can recover alternating hindlimb stepping on a moving treadmill under appropriate conditions. The result demonstrates that spinal networks contain substantial locomotor competence. It does not show that ordinary walking in an intact cat is produced by a sealed spinal clock. Treadmill motion, limb loading, hip position, cutaneous contact, neuromodulatory state, training, and the recovering excitability of the cord all shape the pattern. Removing proprioceptive feedback degrades fundamental aspects of interjoint and flexor–extensor coordination. Cutaneous stimulation at one location can facilitate stepping, while stimulation elsewhere can interrupt it. The spinal generator and the moving body form one coupled system [@bouyerrossignol2003; @rossignol2004recovery].
The human evidence is necessarily less direct. Tonic epidural stimulation of the lumbar cord in people with longstanding, clinically complete spinal cord injury can evoke alternating locomotor-like electromyographic activity. Leg vibration and other sensory manipulations can also elicit stepping-like patterns. These findings support a human spinal capacity for pattern generation. They do not establish how much of ordinary bipedal walking is generated by the same architecture, nor do they justify treating human gait as a spinal program merely started by cortex. Walking over uneven ground, changing direction, carrying a load, or stepping around another person requires continuous cooperation among spinal, brainstem, cerebellar, cortical, sensory, and mechanical systems [@dimitrijevic1998human; @grillnerelmanira2020].
The spinal cord is also a controller outside rhythmic movement. Step on a sharp tack with one foot and the stimulated leg can flex away while extensor activity in the other leg helps support the body. The familiar withdrawal response is therefore not a single flexor wire; it is a coordinated pattern distributed across motor pools. Stretch-related pathways adjust muscle activity as loads change. Cutaneous reflexes are reorganized across the step cycle: an input that assists foot clearance during swing may have a different effect during stance, when the same limb must bear weight. Reflex gain changes with task, posture, expectation, and descending state. Calling these responses reflexes does not mean they are fixed, isolated, or unintelligent. It means that important transformations occur locally and rapidly [@grillnerelmanira2020].
30.5 Evolution adds control above control
The persistence of movement after spinal transection can be misread in two opposite ways. One is to deny the result by treating the cord as a passive cable. The other is to conclude that the brain contributes little beyond a start signal. Both miss what a hierarchy contributes.
Descending systems do not merely turn spinal circuits on. They alter which circuits are recruitable, set their excitability, change reflex gain, coordinate the limbs with the head and trunk, and adapt a recurring pattern to terrain and purpose. The same spinal input can produce different outputs depending on whether an animal is standing, walking, running, scratching, or preparing to move. Higher control is visible precisely because the lower controller has its own dynamics to be governed.
The mesencephalic locomotor region (MLR) provides a particularly clear example. In lamprey and in decerebrate mammalian preparations, graded activation of the MLR recruits reticulospinal systems that initiate locomotion and regulate its speed. The descending signal does not specify every flexor and extensor burst. It changes the drive and operating state of spinal networks whose internal circuitry produces the detailed rhythm. Sensory feedback then adjusts the pattern to loading and terrain. A higher controller can therefore alter whether locomotion occurs and how rapidly it proceeds by parameterizing lower controllers rather than replacing their computation [@grillnerelmanira2020].
Spinal cord injury supplies a revealing example. Immediately after a severe acute injury, reflexes below the lesion are often depressed or absent, muscles are hypotonic, and voluntary movement is lost. This early state is spinal shock. Over days and weeks, reflexes return. They may then become exaggerated: tendon jerks grow brisk, muscle tone increases, clonus and spasms emerge, and small sensory events can recruit large responses. The later hyperreflexia is not simply the restoration of normal spinal function. It reflects the loss of descending modulation together with changes within the isolated cord, including altered synaptic efficacy and increased intrinsic motoneuron excitability [@ko1999spinalshock; @little1999hyperreflexia].
The sequence matters. Cutting descending pathways does not instantaneously reveal an unchanged reflex machine that was waiting underneath. The injury abruptly removes tonic excitation as well as inhibition, producing shock; the cord then reorganizes in the absence of normal supraspinal control. The eventual exaggeration of reflexes nevertheless makes a central point: intact descending systems ordinarily restrain, bias, and contextualize spinal responses. A higher level of control can be demonstrated by what the lower level does when that control is lost.
This is hierarchy without a homunculus. No single descending neuron decides the movement and specifies every contraction. Brainstem and cortical systems project through broad and partly distinct channels. Some are especially important for balance, axial musculature, orienting, and locomotion. Others provide more direct access to distal muscles and fractionated hand movements. They converge on spinal interneurons and motoneurons already participating in local feedback loops. A voluntary action is therefore an intervention into an active system, not a command sent to dormant hardware.
The evolutionary interpretation follows naturally. Older circuits generated useful patterns before later vertebrates acquired the enlarged pallial systems of mammals. New systems increased the range of conditions under which those patterns could be selected, combined, redirected, or suppressed. They also created forms of control that are difficult to reduce to a ready-made program: shaping a hand to an unfamiliar object, playing a newly learned sequence, imitating a gesture, or adjusting a reach according to an abstract rule. The older hierarchy remained, but the space of actions available to it expanded.
30.6 The body is part of the controller
A nervous system does not control abstract points moving through empty space. It controls a body with mass, elasticity, friction, joint limits, tendons, muscles, and tools. Those mechanics are not an inconvenient final stage after the neural computation. They help determine what control is required and what control is possible.
Consider a rapid elbow movement. Accelerating the forearm produces forces at the shoulder. Rotating the shoulder changes the torques required at the elbow. The amount of force generated by a muscle depends on its length, recent activity, and shortening velocity. Tendons store and return elastic energy. The same neural drive produces different movement when the arm carries a suitcase, moves through water, or rests against a surface. A command specified only as “activate the biceps” has no fixed consequence outside the current mechanical state of the limb.
The body can also simplify control. Tendon elasticity smooths force and returns energy during locomotion. The geometry of joints constrains possible trajectories. Passive dynamics can carry a limb through part of a step. Co-contraction can increase stiffness when uncertainty makes flexibility dangerous. The nervous system sometimes works against body mechanics and sometimes recruits them. Skilled action depends on learning which is which.
This is why proprioception is not a sensory appendix to movement. Muscle spindles, tendon organs, joint-related signals, cutaneous receptors, vestibular organs, and vision report different aspects of the moving body. Their signals are delayed, noisy, and sometimes inconsistent, but without them the controller rapidly loses accuracy and stability. The fastest peripheral afferents are devoted largely to proprioception because an estimate of the body that arrives too late is of limited use for a limb already in motion.
The body and nervous system should therefore be treated as a neuromechanical system. Neural activity changes force; force changes posture and contact; those changes alter sensory input; and the next neural state emerges from the entire loop. In engineering language, the body is the plant being controlled. In biological language, the plant is not passive. Its structure is one of the mechanisms by which control succeeds.
30.7 Getting force to muscle
Whatever its origin, neural control reaches skeletal muscle through lower motor neurons in the spinal cord and cranial motor nuclei. Sherrington called them the final common pathway. The phrase remains useful because it states a hard anatomical constraint. A cortical plan, a brainstem postural response, a spinal reflex, and a pattern generator can all converge on the same motoneuron pool. If the relevant lower motor neurons or their axons are destroyed, the muscle loses its ordinary neural route to contraction.
A lower motor neuron and the muscle fibers it innervates form a motor unit. Force can be graded by recruiting additional motor units and by changing their firing rates. Small, fatigue-resistant units are generally recruited before larger, more forceful ones, allowing smooth control across a wide range. Motoneurons are not simple relays. Their excitability is altered by thousands of synaptic inputs, neuromodulators, recurrent inhibition, and intrinsic conductances that can sustain firing. Even at the final pathway, output depends on state.
Above them lies an extensive spinal interneuronal network. Some interneurons link sensory afferents to motor pools; others coordinate muscles across joints and sides of the body; still others carry descending influences into local circuitry. The spinal cord is where descending goals meet the current state of the limb. That convergence helps explain why the same cortical activity can have different consequences when posture, load, or phase changes.
Descending control arrives from both cerebral cortex and brainstem. Brainstem pathways contribute strongly to posture, balance, head and body orientation, and control of axial and proximal musculature. The corticospinal tract is especially important for flexible voluntary action and for the fractionated control of distal muscles, particularly the primate hand. Some corticospinal neurons influence motoneurons through spinal interneurons; a subset in primates makes direct cortico-motoneuronal connections, providing unusually direct cortical access to hand and finger muscles [@rathelotstrick2006; @rathelotstrick2009].
The familiar distinction between medial systems for axial and proximal control and lateral systems for distal control is useful, but it is a bias rather than a clean separation. Reaching requires trunk stabilization, shoulder transport, elbow coordination, wrist orientation, and finger shaping. Brainstem and cortical pathways cooperate across those components. Skilled action does not begin only when the signal reaches the hand.
The crossing of descending pathways also requires a calibrated account. Most corticospinal axons cross in the lower medulla, and unilateral cortical damage commonly causes predominantly contralateral weakness, especially for skilled distal movements. But the system is not simply one hemisphere controlling only the opposite half of the body. Some fibers descend uncrossed, some cross within the spinal cord, and many projections terminate bilaterally or influence bilateral interneuronal networks. Axial and proximal control is particularly bilateral. The contralateral bias is strong and clinically consequential; the underlying wiring is more distributed than the motor homunculus suggests [@rosenzweig2009].
30.7.1 What is represented in motor cortex?
Primary motor cortex contains a coarse somatotopic organization: leg-related territory lies medially, face-related territory laterally, and arm and hand territory between them. The maps overlap, repeat, and vary with the method used to reveal them. Individual muscles are not represented as isolated keys on a keyboard. Cortico-motoneuronal cells influencing one finger muscle can be distributed broadly and intermingled with cells influencing others.
Long trains of cortical microstimulation in monkeys can evoke coordinated, multijoint movements that bring the hand toward the mouth, shape the hand, or orient the body into an action-like posture. These findings helped correct the image of a cortex divided into tiny single-muscle switches. They do not prove that each site stores one complete ethological action. Electrical stimulation imposes an artificial pattern on local and connected tissue, and what a circuit can be driven to produce is not identical to what it normally represents [@graziano2002complex].
Recordings during voluntary movement support several descriptions at once. Individual neurons can covary with muscle activity, force, direction, posture, movement phase, and sensory feedback. Population activity evolves through structured dynamics that can generate temporally patterned output. These are not mutually exclusive findings. A neural population can have lawful internal dynamics while its output remains related to muscles and forces. The open question is which description best explains the system across tasks, not whether one vocabulary must defeat all the others [@churchland2012dynamics; @russo2018].
The motor homunculus is therefore useful as a first anatomical orientation and dangerous as a theory. There is no little body laid out in cortex and no smaller agent moving it. There are overlapping cortical populations embedded in loops with parietal, premotor, somatosensory, basal-ganglia, cerebellar, thalamic, brainstem, and spinal systems. Their activity gains meaning through those connections and through the body they control.
30.8 Acting on the seen world
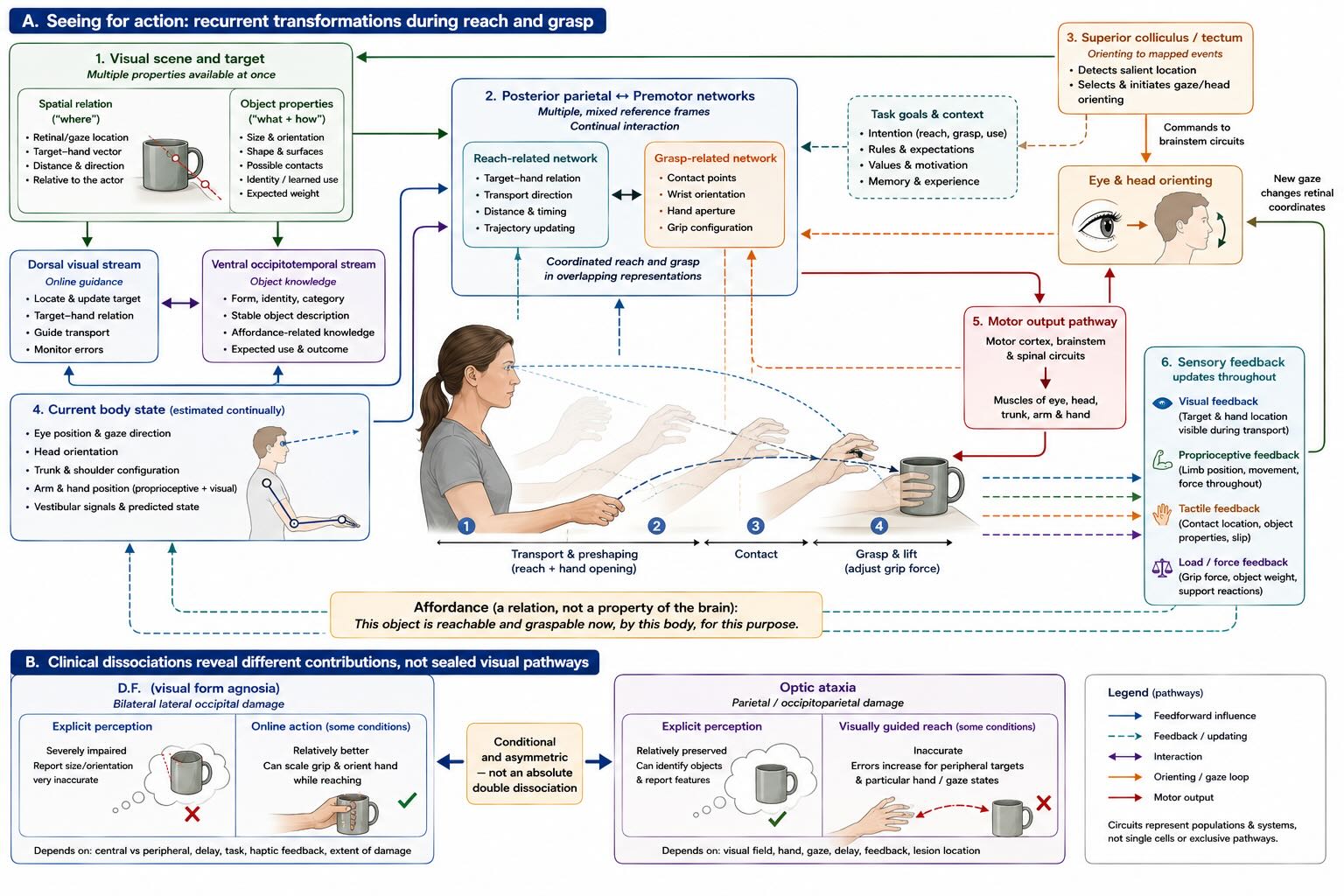
The unit on sensation repeatedly asked what sensory information is for. Visually guided action gives the direct answer. Seeing an object is not enough. To reach for it, the nervous system must relate the object’s location to the eyes, head, trunk, shoulder, hand, and possible movement. To grasp it, the system must estimate size, orientation, shape, surface, and load while deciding where the fingers can make contact. The action changes the view, the arm configuration, and the tactile evidence, so the estimate must be updated while movement unfolds.
No single coordinate system is sufficient. Retinal location is initially defined relative to gaze. The hand is represented through proprioceptive and visual signals in relation to the shoulder and body. The target may move, the eyes may move, and the head may turn before the reach is complete. Posterior parietal and premotor networks contain populations sensitive to overlapping combinations of gaze, target, hand, and body state. Rather than performing one final conversion from “visual coordinates” into “motor coordinates,” these networks appear to maintain several partial reference frames whose relative influence depends on the task.
The dorsal visual networks introduced in Unit IV contribute especially to online guidance: locating a target relative to the actor, updating that location, shaping the hand, and correcting a trajectory as conditions change. Ventral visual networks contribute more strongly to stable descriptions used for recognition, memory, and object-centered knowledge. The division is real enough to generate striking dissociations and interactive enough that it should never be drawn as two sealed pipes [@goodalemilner1992; @goodalewestwood2004].
Patient D.F., with severe visual form agnosia after bilateral damage centered on lateral occipital regions of the ventral visual stream, provided the classic example. She had great difficulty reporting an object’s size or orientation yet could, under some conditions, scale her grip and orient her hand while reaching toward it. Patients with optic ataxia after parietal damage can show the complementary problem: relatively preserved perception with inaccurate visually guided reaching, especially toward peripheral targets. These cases demonstrate that information supporting explicit perceptual report can dissociate from information supporting online action [@goodale1991; @milner1991agnosia; @pereninvighetto1988].
They do not establish complete independence. D.F.’s performance depends on task conditions, central versus peripheral presentation, available haptic calibration, delay, and the extent of damage beyond the ventral stream. Patients with optic ataxia can improve when a target is foveated or when additional time permits other representations to contribute. Recognizing what an object is also affects how it is grasped for use. The dorsal and ventral systems make different contributions within a connected visual–motor network [@schenk2012; @goodalewestwood2004].
The evolutionary hierarchy is visible here too. The optic tectum of nonmammalian vertebrates and its mammalian homologue, the superior colliculus, link mapped sensory events to orienting of the eyes, head, and body. Expanding parietal and frontal systems did not make this midbrain machinery obsolete. They added more flexible reaching, grasping, rule use, delayed action, and tool use while continuing to interact with the tectum, basal ganglia, cerebellum, thalamus, and brainstem.
The relevant unit of visual control is therefore not a picture transmitted to a motor center. It is an affordance relative to the current body: something reachable now, graspable with this hand, avoidable along this trajectory, or worth orienting toward given the animal’s state. The term does not imply that perception is infallible or that all object knowledge is motoric. It keeps the geometry of action where it belongs—between an embodied agent and a structured world.
30.9 Posture is active stabilization
Reaching begins before the arm moves. A standing body must control its relation to gravity and the support surface while the movement changes forces throughout the body. Even apparent stillness is a dynamic achievement. The center of pressure shifts beneath the feet, muscles alter their activity, vestibular and proprioceptive signals change, and the body sways within a small region without falling.
It is tempting to call posture the defense of a set point. That analogy captures the importance of feedback but is too literal if it suggests one fixed ideal pose. Postural control stabilizes different variables under different conditions. It may keep the center of mass within the base of support, the head oriented relative to gravity, gaze on a target, a hand steady in external space, or a tray level while the person walks. The defended reference changes with task, and the acceptable region may be broad rather than a single point.
Posture is therefore better described as task-dependent stabilization. The nervous system allows movement that does not threaten the task and counters movement that does. When the support surface narrows, the available solutions change. When a person holds a rail, the hand becomes part of the postural system. When the head must remain stable for vision, neck and trunk muscles are coordinated around that requirement. Balance is not one reflex; it is the coordinated regulation of several variables through sensory and motor systems distributed from spinal cord to cortex.
30.10 Prediction and correction are one control process
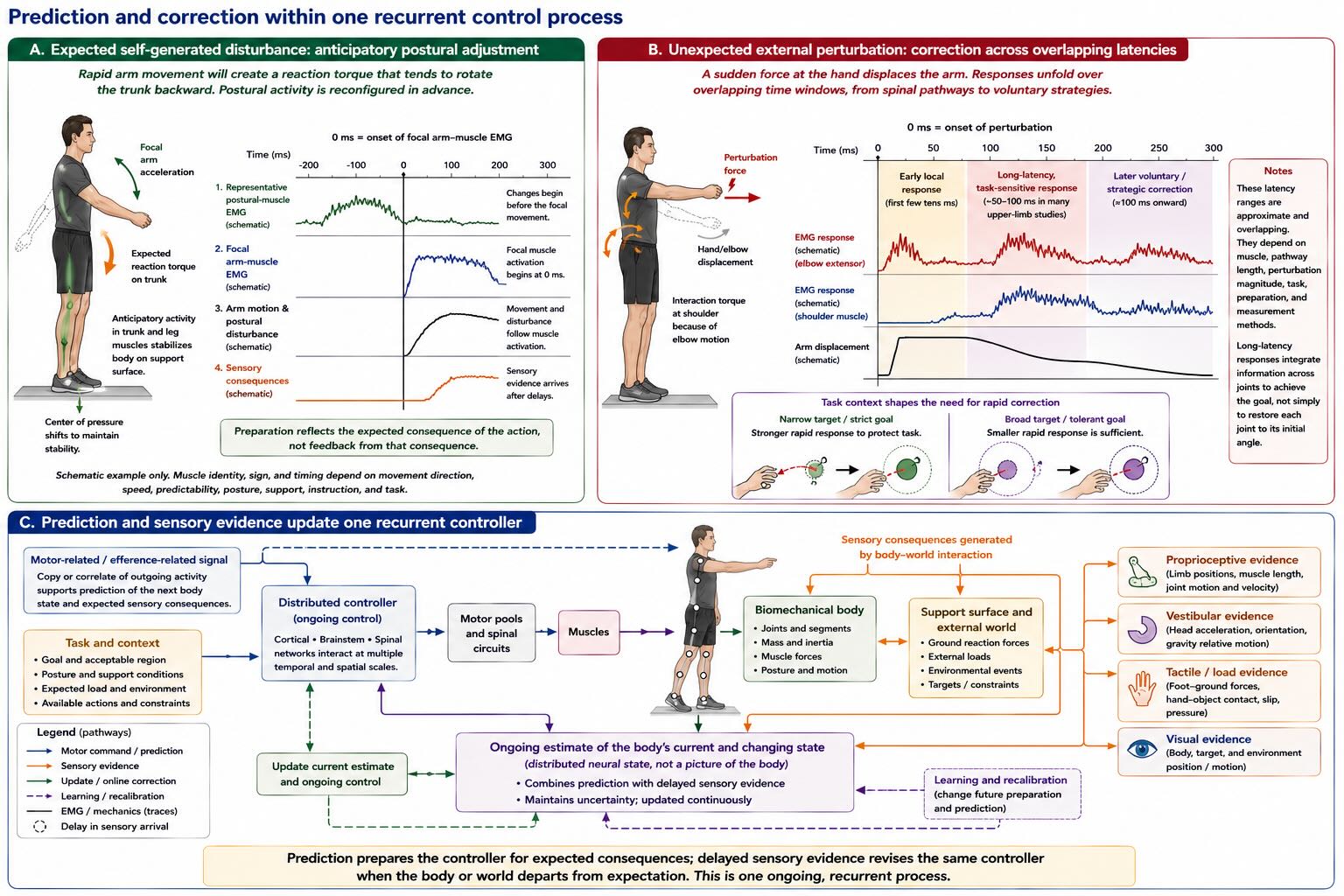
Purely delayed correction would be a poor strategy for a body. Neural conduction, receptor transduction, muscle activation, and the mechanics of movement all take time. By the time sensory evidence reports that a rapid movement has destabilized the trunk, the disturbance is already underway. The motor system therefore prepares for predictable consequences.
An anticipatory postural adjustment makes the principle visible. Before a rapid arm movement, activity can change in leg, trunk, and even respiratory muscles. The exact muscles, signs, and timing depend on movement direction, support conditions, speed, predictability, and instruction. There is no universal rule that one calf muscle always fires before one arm muscle. The robust finding is that the postural system can be reconfigured before the focal movement produces its mechanical disturbance [@cordonashner1982].
This preparation is often called feedforward control, whereas responses driven by incoming sensory evidence are called feedback control. The distinction is useful as long as it does not become a division into two separate machines. Anticipatory commands are built from prior feedback and continue to be revised by feedback. Feedback responses are not generic corrections applied after the plan is complete. Their gain and direction depend on the current goal, posture, expected load, and available alternatives.
An unexpected perturbation to the arm produces a sequence of responses. Very short-latency activity reflects local spinal pathways and the current state of the motoneuron pool. Longer-latency activity can incorporate information across joints and is sensitive to the behavioral goal. Later voluntary corrections can use still wider information and change strategy. The boundaries overlap. A response can be rapid and intelligent because higher systems have already configured the pathways through which the incoming signal will act.
Control also requires an estimate of the body’s current state. Sensory signals arrive after different delays and contain different uncertainties. Vision may accurately report hand position but be temporarily unavailable; proprioception is fast but can be biased; an efference copy of outgoing motor activity can support a prediction of consequences before reafferent input arrives. A state estimate combines these sources to infer where the body is and how it is changing now. The estimate need not be a little internal picture. It is the pattern of activity that allows the next control signal to be appropriate to a body whose state cannot be sampled instantaneously.
The same logic explains rapid multijoint feedback. A perturbation at the elbow changes shoulder forces because the joints are mechanically coupled. An effective response cannot treat each joint independently. Primary motor cortex participates in fast feedback that incorporates these interaction torques, showing again that cortex is not only the origin of voluntary commands. It is part of a recurrent controller responding to consequences while movement unfolds [@pruszynski2011feedback].
Prediction and feedback are therefore complementary aspects of one process. Prediction prepares the system for expected consequences and partly compensates for delay. Feedback reveals what actually happened and corrects the estimate, the current action, and future preparation. Learning changes both.
Lift two objects that have the same mass but very different sizes. The smaller object usually feels heavier. This size–weight illusion persists even after many alternating lifts.
The forces used to lift the objects tell a different story. On the first encounter, people typically prepare more force for the larger object because experience has taught them that larger objects of similar material usually weigh more. After several lifts, grip and load forces adapt to the objects’ actual equal masses. The sensorimotor prediction becomes accurate while the perceptual illusion remains [@flanaganbeltzner2000].
The result is important because it separates two claims that are easy to combine. It shows that perceptual judgment and the force predictions used for lifting can be updated partly independently. It does not, by itself, identify one anatomical “motor model,” prove that the motor estimate is globally more accurate than perception, or demonstrate that the cerebellum alone produced the adaptation. It establishes a dissociation that any adequate theory of prediction must explain.
30.11 The cerebellum: calibration in a delayed system
The cerebellum is positioned to address the problems just described. It receives extensive information about ongoing motor commands, body state, and sensory consequences. Its outputs return through brainstem and thalamic routes to systems controlling movement. Its circuitry is remarkably regular, and related architectures are conserved across vertebrates. Yet a compact description of what the cerebellum does remains difficult.
Several functions are secure. Cerebellar damage produces ataxia: movements become poorly timed, inaccurately scaled, decomposed, or unstable. Reaching can overshoot or undershoot. Rapid alternating movements become irregular. Balance and eye movements are disturbed. The deficit is not ordinarily paralysis; the muscles can produce force, but the force is not coordinated with sufficient accuracy across time and effectors.
The cerebellum is also essential for many forms of error-based adaptation. When a visual display shifts the apparent consequence of a reach, healthy participants gradually alter their commands. People with cerebellar degeneration adapt less. Experiments that separate online correction from trial-to-trial learning indicate that the critical teaching signal often behaves like a sensory prediction error: a mismatch between the sensory consequence expected from a command and the consequence observed [@tseng2007cerebellum; @shadmehretal2010].
These findings motivate the influential proposal that the cerebellum learns internal models. A forward model predicts the sensory consequences of a motor command; an inverse model estimates commands capable of producing a desired result. Predictions can help compensate for delay, coordinate effectors, and distinguish expected reafference from an unexpected disturbance. The framework unifies a large body of behavior and physiology [@wolpertmiall1996; @shadmehretal2010].
It should remain a framework rather than a definition. Timing, state estimation, calibration, coordination, and prediction may describe related functions at different levels, and no single internal-model formulation yet explains every cerebellar contribution. The cerebellum also participates in cognitive, affective, autonomic, and language-related loops whose relation to motor prediction remains actively studied. Calling it a “prediction machine” is productive when it generates tests and misleading when it closes the question.
The chapter The Predicting Machine will therefore treat prediction as a strong hypothesis with unusually good motor evidence, not as a slogan that makes all cerebellar functions settled.
30.12 The basal ganglia: choosing among available actions
A nervous system with many competent action systems has a new problem. It cannot orient toward every stimulus, approach and withdraw at once, or release every motor pattern made available by the spinal cord and brainstem. Some actions are incompatible; others must be sequenced. Selection is therefore not an optional cognitive layer added after movement. It is a basic requirement of a system containing competing controllers.
The basal ganglia are central to this problem. Cortical or pallial and thalamic inputs converge on the striatum, while basal-ganglia loops also interact extensively with tectal and other brainstem systems. Basal-ganglia output nuclei maintain strong inhibitory influence over thalamic and brainstem targets. Activity through several pathways can reduce inhibition for selected channels, increase it for competitors, regulate the speed and vigor of an action, and alter which transitions are likely next. The architecture is often summarized as a gate, but the gate is distributed, channelized, and embedded in recurrent loops.
The familiar direct-pathway “go” and indirect-pathway “no-go” diagram is a useful first scaffold. It captures the opposing signs of important routes through striatum, pallidum, subthalamic nucleus, and output nuclei. It becomes inaccurate when treated as two populations that take turns switching all movement on or off. Direct- and indirect-pathway neurons can be active together around action initiation. Their relative timing and spatial pattern may help facilitate selected components while restraining alternatives, adjust vigor, and shape learning. The hyperdirect cortical route through the subthalamic nucleus adds another way to influence output rapidly. Selection is not one lever [@cui2013concurrent; @yttridudman2016].
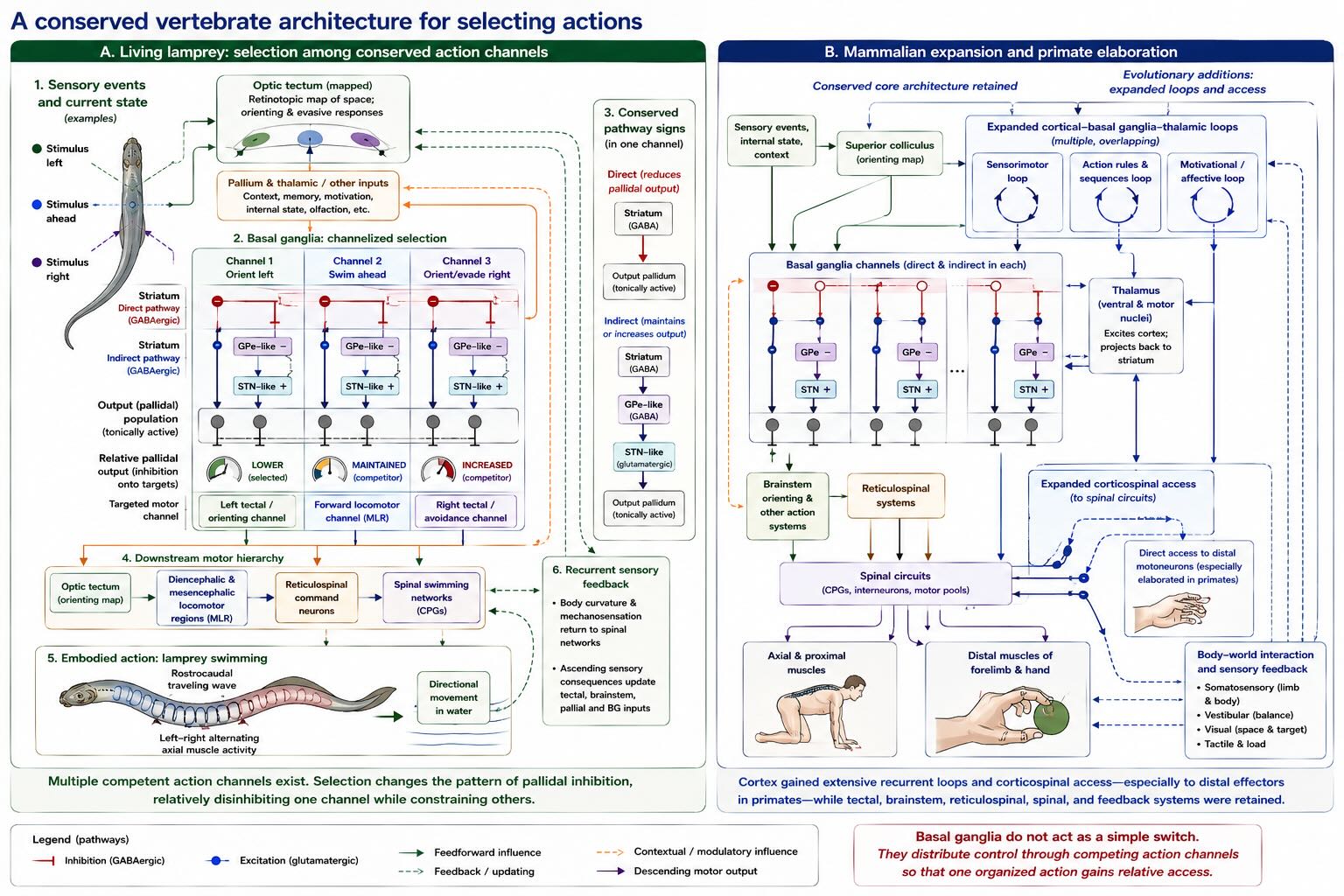
30.12.1 Why the lamprey clarifies the problem
The lamprey is particularly useful because the relevant motor hierarchy can be studied across levels that are difficult to connect in a mammal. Its spinal cord contains well-characterized swimming networks. Brainstem systems can initiate and steer locomotion. The optic tectum maps sensory space and contributes to orienting. Its forebrain contains a basal-ganglia architecture recognizable in molecular markers, cell types, direct and indirect pathways, subthalamic circuitry, and tonically active pallidal output [@stephensonjones2011basal; @ericsson2011striatal; @grillnerrobertson2016].
Sten Grillner and his colleagues used this preparation to make an evolutionary argument unusually concrete. Pallidal output in lamprey tonically inhibits tectal and other motor-related regions. Striatal activity can alter that inhibition so that one action system is released while others remain constrained. The basal ganglia need not specify each left–right bend of the swimming body. Spinal networks already know how to generate the pattern. The higher system helps determine whether this pattern, in this direction, under these conditions gains access to the downstream machinery [@stephensonjones2011basal; @grillnerrobertson2016].
Lampreys belong to an early-diverging vertebrate lineage, but living lampreys are not preserved ancestors. Their lineage has continued to evolve for more than half a billion years. They are informative because many components of the vertebrate motor plan are accessible and because comparisons reveal which features were already present near the base of vertebrate evolution [@grillner2021evolution; @ocana2015pallium].
The important conclusion is not that a human action is secretly a lamprey swim with cortex placed on top. It is that the ancestral vertebrate already possessed a layered architecture in which basal-ganglia output influenced tectal and brainstem motor systems, which in turn governed spinal pattern-generating circuitry. Mammalian evolution expanded the cortex, multiplied cortico-basal-ganglia-thalamic loops, increased direct cortical access to motoneurons, and enlarged the repertoire of learned and abstract actions. It retained the core logic of selection through a conserved subcortical system.
Comparative work is especially valuable here because the human basal ganglia participate in motor, cognitive, emotional, and motivational loops that can make their common architecture hard to see. The lamprey does not reduce those human functions to locomotion. It reveals the older control problem from which the expanded loops could be elaborated.
This perspective also improves the interpretation of disease. Parkinson’s disease is not simply a closed motor gate, and Huntington’s disease is not simply an open one. Parkinsonian bradykinesia, rigidity, tremor, impaired initiation, and reduced spontaneous movement arise from altered dynamics across basal-ganglia, cortical, thalamic, and brainstem networks following degeneration of dopamine neurons and other changes. Huntington’s disease produces involuntary movement but also impairs voluntary action, cognition, and emotion as different striatal and cortical populations degenerate. The disorders reveal the importance of selection, scaling, and sequencing without reducing the basal ganglia to a binary switch.
30.13 What an action is worth
Selection requires criteria. A hungry animal should approach food that a sated animal may ignore. A route that led to injury should become less likely. An action that produced an unexpectedly good outcome should gain influence when similar conditions return. The motor hierarchy therefore intersects with systems for motivation, valuation, and learning.
Dopamine is central to that intersection, but it is not a chemical synonym for reward or pleasure. In classic conditioning experiments, many midbrain dopamine neurons respond strongly to an unexpected reward. As a predictive cue is learned, the response shifts toward the cue; omission of the expected reward produces a dip around the time it should have occurred. This pattern resembles a reward-prediction error: information about whether an outcome was better or worse than expected. Such a signal can modify recently active corticostriatal synapses and thereby alter future action selection [@schultzdayanmontague1997].
That account is powerful and incomplete. Dopamine neurons and their axons are heterogeneous. Signals differ across projection targets. Dopamine activity can covary with locomotion, movement initiation, salience, motivational state, and the vigor of future action as well as with reward prediction errors. Local striatal circuitry further controls when and where dopamine is released. A single scalar “value signal” is therefore an introductory approximation, not the whole biology [@panigrahi2015vigor; @howedombeck2016; @dasilva2018dopamine].
The distinction between wanting and liking makes a related point. Cues can acquire excessive motivational pull without producing a correspondingly intense pleasurable experience. That dissociation helps explain why compulsive pursuit can persist after enjoyment has weakened. It does not mean addiction is a lesion of one wanting center. Addiction changes distributed systems for learning, motivation, stress, habit, choice, and control, and those changes interact with drug pharmacology and the person’s social environment [@berridgerobinson1998].
The final chapter of the unit, What It’s Worth, will ask how outcomes alter future actions. The question belongs beside motor control because movement is not merely executed; it is continually reweighted by what its consequences did to the organism.
30.14 What failure reveals
Clinical syndromes are useful because they separate control problems that ordinary movement hides. They should not be treated as a lookup table in which one structure equals one deficit. Real lesions cross pathways, disconnect regions, and alter networks over time. Even so, several profiles provide a durable orientation.
| Principal disruption | Typical consequences | What the profile suggests |
|---|---|---|
| Lower motor neuron or peripheral motor axon | Weakness or paralysis, muscle atrophy, reduced tone and reflexes | The final neural route to a muscle has been damaged |
| Motor cortex or corticospinal pathways | Predominantly contralateral weakness, loss of fractionated movement, and—after the acute phase—abnormal tone and reflexes | Flexible voluntary access and descending regulation of spinal circuitry have been impaired |
| Spinal sensory guidance or severe deafferentation | Poorly controlled movement that becomes highly dependent on vision | Force alone is insufficient; action requires continuous estimates of body state |
| Posterior parietal visuomotor networks | Optic ataxia and impaired online reaching under characteristic conditions | Visual perception and visual guidance of action can dissociate |
| Distributed left praxis network | Limb apraxia: impaired learned skilled action not explained by elementary weakness | Action organization can fail above basic force production |
| Cerebellar circuits | Ataxia, dysmetria, decomposition, timing deficits, impaired adaptation | Calibration and coordination can fail while strength remains relatively preserved |
| Basal-ganglia and dopamine systems | Bradykinesia, rigidity, tremor, involuntary movement, altered vigor, habit, and learning | Selection and scaling depend on recurrent subcortical loops, not cortex alone |
The distinction between cortical weakness and apraxia is especially important. Damage to primary motor cortex or the corticospinal tract commonly produces weakness and loss of fine control. Apraxia is a higher-order impairment of learned skilled action that cannot be explained adequately by weakness, sensory loss, or incomprehension. It is most often associated with a distributed left-hemisphere network involving parietal, frontal, temporal, and connecting white-matter systems. A patient may have enough strength to move the hand yet fail to organize a familiar gesture correctly. Calling all cortical motor impairment apraxia erases the distinction the syndrome was created to mark [@kusch2018apraxia].
Spinal injury provides a complementary lesson. Early areflexia, later hyperreflexia, involuntary spasms, and the occasional recovery of stepping-like patterns arise from different processes below and above the lesion. No single symptom tells us that the cord is either passive or autonomous. The profile changes as excitability and circuitry change.
The general rule is to ask what control problem has failed: generating force, stabilizing posture, estimating body state, using vision online, organizing learned action, calibrating prediction, selecting among alternatives, or updating action from outcome. Anatomical localization then becomes more accurate because it is constrained by the full pattern rather than by one dramatic symptom.
30.15 How motor control is studied
Motor neuroscience draws evidence from preparations that expose very different parts of the hierarchy. Understanding what each method removes is as important as knowing what it records.
Reduced and isolated preparations reveal intrinsic circuit capacity. Fictive locomotion shows that patterned motor output can persist without movement-generated sensory feedback. Spinalized animals show what cord and peripheral feedback can accomplish without normal supraspinal input. These experiments are indispensable for identifying CPGs and reflex organization. They do not reproduce the intact behavior of an uninjured, freely moving animal, and the conditions used to activate the circuit—drugs, electrical stimulation, treadmill loading, or tonic drive—must be part of the interpretation.
Kinematics, force, and electromyography reveal how the body moves and which muscles are active. They can expose anticipatory adjustments, synergies, adaptation, and the timing of correction. EMG does not identify the neural source of a response by itself. Similar muscle patterns can be produced through different combinations of spinal, brainstem, and cortical activity.
Mechanical and sensory perturbations are especially informative because they force the controller to reveal what it is stabilizing. Moving the target, pushing a joint, changing a load, shifting visual feedback, or altering the support surface separates prediction from correction and task-relevant from task-irrelevant error. A perturbation experiment often tells more about control than observation of an unperturbed reach, because many candidate mechanisms produce the same successful trajectory until the environment pushes them apart.
Electrical or magnetic stimulation establishes causal access. If stimulation of a cortical territory evokes a coordinated movement, that tissue can influence the movement through the recruited network. The result does not show that the normal code consists of the movement evoked, especially when stimulation is prolonged or synchronizes neurons that would not normally fire together.
Single-neuron recordings and population analyses answer related but different questions. A neuron may covary with direction, force, muscle activity, or feedback. A population may display rotational or other dynamical structure that is not obvious in single cells. Population dynamics do not make the relationship to muscle irrelevant, and muscle correlations do not show that each neuron is an independent command line. A useful theory must explain how internal population activity produces output appropriate to the body and task.
Lesions and disease establish necessity more directly than activation alone, but lesions rarely respect functional borders. Vascular territories, white-matter disconnections, diaschisis, compensation, medication, and time since injury all matter. Patient D.F., optic ataxia, apraxia, ataxia, Parkinson’s disease, and spinal injury are most informative when the behavioral profile, anatomy, and converging physiology are considered together.
Comparative anatomy and physiology reveal the evolutionary architecture. Similar cell types, pathways, molecular markers, and circuit signs across lamprey and mammals provide stronger evidence of conservation than superficial behavioral resemblance alone. The comparison must not turn living species into rungs on a ladder. Each lineage is specialized; the goal is to infer which components were inherited and how their relative influence changed.
Finally, computational models make proposed control principles explicit. Internal models, optimal feedback control, dynamical systems, reinforcement learning, and action-selection models can unify observations and generate quantitative predictions. They are explanations at a chosen level, not automatically literal diagrams of the tissue. A model earns biological weight when its assumptions survive perturbation, lesion, physiology, and comparative evidence.
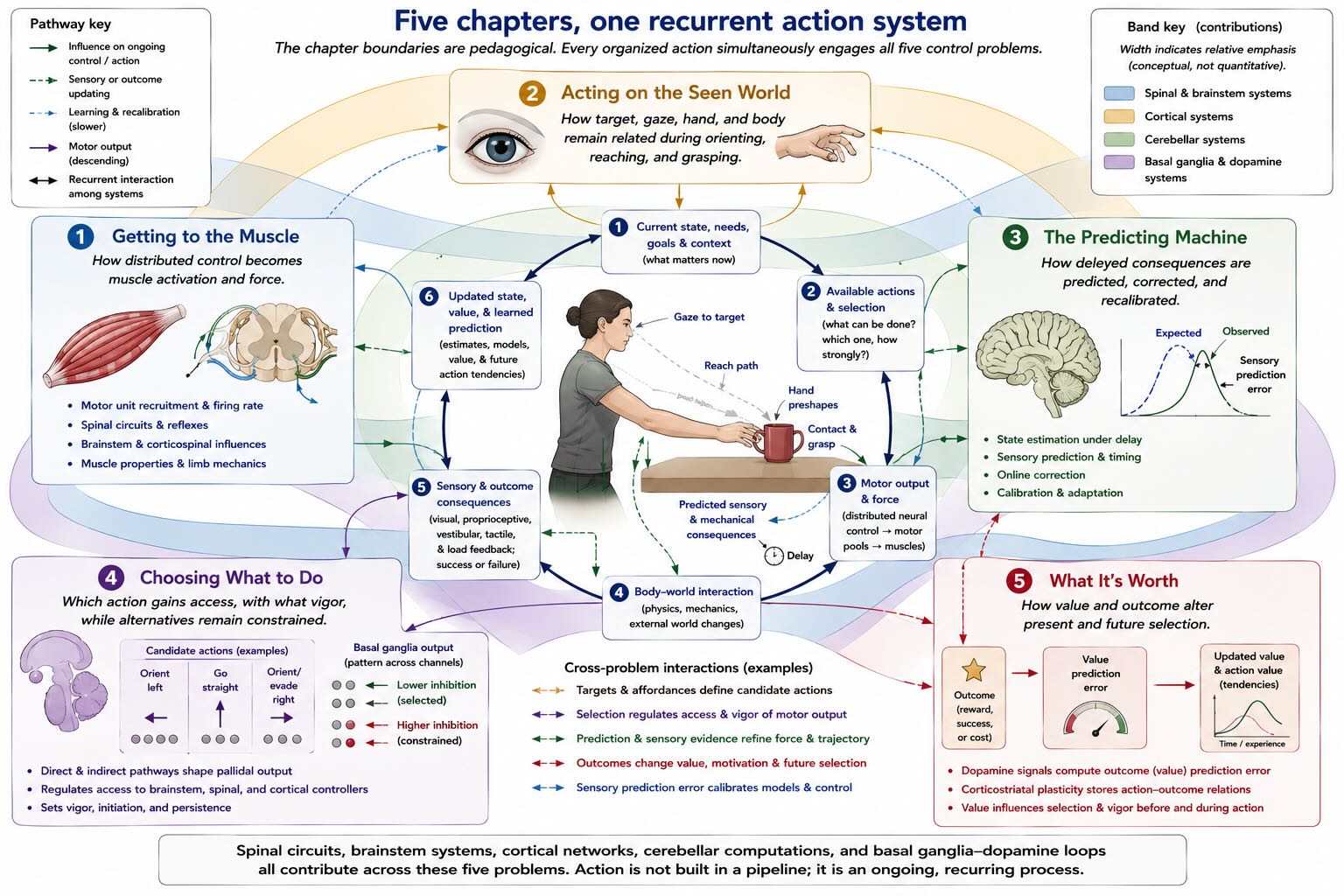
30.16 The five problems ahead
This unit divides one recurrent system into five chapters because no single chapter can develop every control problem at once. The divisions are pedagogical, not anatomical stages through which a command passes in order.
Getting to the Muscle begins with the final common pathway and works upward through motor units, spinal circuitry, brainstem systems, corticospinal control, motor cortex, and the clinical distinction between lower- and upper-motor-neuron disorders. Its central question is how force becomes available to an action without requiring cortex to specify every contraction.
Acting on the Seen World asks how visual and body-centered information guide orienting, reaching, and grasping. It develops coordinate transformations, posterior parietal and premotor networks, dorsal–ventral interactions, and the dissociations revealed by visual form agnosia and optic ataxia.
The Predicting Machine turns to the cerebellum and to the problem of delay. It asks how a controller estimates current state, anticipates sensory consequences, calibrates movement from error, and learns when the relation between command and consequence changes.
Choosing What to Do develops the basal ganglia as an ancient system for influencing which actions gain access to downstream controllers, how strongly they are expressed, and which alternatives remain suppressed. The lamprey provides the evolutionary scaffold; Parkinson’s and Huntington’s diseases show what happens when the expanded mammalian network fails.
What It’s Worth asks how consequences alter later selection. Dopamine, corticostriatal plasticity, prediction error, incentive motivation, habit, and addiction enter here. Value is not added after movement. It is one of the variables that determine which movement becomes action.
The chapters will repeatedly cross their assigned borders. Cerebellar feedback enters cortical control; basal ganglia influence brainstem systems as well as cortex; sensory pathways reach spinal circuits before perception; dopamine changes motor vigor as well as learning. The organization is therefore best read as five views of one hierarchy.
30.17 Coda: control becomes consequence
A bacterium can bias a random walk without neurons. A spinal network can generate a rhythm without a patterned command from the brain. A lamprey basal ganglion can help release one orienting or locomotor system while constraining another. A primate cortex can shape individual fingers around a tool, but it does so through inherited brainstem and spinal machinery, a biomechanical hand, and sensory feedback that begins correcting the action before awareness catches up.
The continuity is not sameness. A human action system supports delayed rules, arbitrary symbols, tools, imitation, speech, music, and social commitments whose immediate connection to survival may be remote. Those capacities depend on enormous expansions and reorganizations of forebrain control. The evolutionary point is that the expansion did not abolish the lower levels. It created more powerful ways to select, parameterize, combine, and learn through them.
Hierarchy is therefore not an embarrassment to be removed from motor theory. It is one of the principal facts to explain. The spinal cord controls muscles and reflex gains. Brainstem systems coordinate posture, orientation, and recurring action patterns. Basal ganglia influence access and vigor. Cerebellar circuits calibrate predictions and corrections. Parietal and frontal cortex add flexible sensory guidance, rules, and unusually direct access to distal effectors. Each level remains recurrently coupled to the body and to the levels around it.
The result is not a puppeteer but an architecture of controllers controlling controllers. Action is what that architecture does when it changes the world and receives the consequences back.
We begin with the point at which every somatic action must finally become force: Getting to the Muscle.
We are confident that:
- Somatic action is produced by distributed, hierarchical, and recurrent interactions among spinal cord, brainstem, cerebral cortex, cerebellum, basal ganglia, sensory systems, muscles, and body mechanics.
- Lower motor neurons provide the final neural route to skeletal muscle, while spinal interneuronal circuits perform consequential local transformations rather than merely relaying commands.
- Spinal and brainstem networks can generate substantial structure in rhythmic and reflexive behavior without moment-to-moment cortical specification. Useful intact behavior nevertheless depends strongly on sensory and descending modulation.
- Severe acute spinal injury often produces an early period of spinal shock, followed later by the return and frequent exaggeration of reflexes as descending control is lost and spinal circuitry changes.
- Motor cortex has a broad somatotopic organization, but representations overlap, direct cortico-motoneuronal access is concentrated in particular primate territories, and cortical activity cannot be reduced to a keyboard of isolated muscles.
- Visually guided action depends strongly on interacting posterior parietal, premotor, motor, subcortical, and sensory systems. Perceptual report and online visual guidance can dissociate without becoming wholly independent.
- Anticipatory postural adjustments, rapid feedback corrections, cerebellum-dependent adaptation, and basal-ganglia contributions to action selection and vigor are well-established phenomena.
- The core basal-ganglia architecture is deeply conserved across vertebrates; lamprey data support an ancient role in influencing access to tectal and brainstem motor systems.
We have good reason to think that:
- Motor abundance is normally exploited so that task-relevant variables are stabilized more strongly than irrelevant details, as proposed by optimal-feedback and related coordination theories.
- State estimation and prediction help compensate for sensory and motor delays, and cerebellar forward-model-like processes contribute importantly to calibration and adaptation.
- Structured motor-cortical population dynamics provide a mechanistic level that complements, rather than simply replaces, descriptions in terms of muscles, forces, and movement variables.
- Basal-ganglia output helps select, scale, sequence, and invigorate actions by modulating competing channels, with direct, indirect, and hyperdirect pathways contributing in more flexible ways than a simple go/no-go switch.
- Dopamine-related prediction errors help modify corticostriatal connections so that recent actions become more or less likely under similar future conditions.
We remain genuinely unsure about:
- How directly the detailed organization of locomotor CPGs established in lamprey, cat, and other preparations maps onto ordinary intact human walking.
- Which description provides the most general account of primary motor cortex across behaviors: muscle and force output, task variables, control policy, population dynamics, or some integration of these levels.
- Whether one internal-model computation can unify the full range of cerebellar motor and nonmotor functions.
- Exactly how basal-ganglia pathways divide labor among selection, suppression, vigor, sequence, habit, and learning across different striatal territories and species.
- How reward-prediction-error, movement-related, motivational, and salience-related dopamine signals are distributed across neurons, axons, projection targets, and timescales.
The productive position is neither that the motor system is a top-down command chain nor that hierarchy has disappeared into a flat network. Evolution retained local controllers, added controllers above them, and connected the levels recurrently through the body and world. The broad architecture is secure. Several of its most interesting computations remain open.