Somatosensation begins at the boundary between the internal and external worlds. It reports events already occurring at the body surface and the configuration of the body itself: pressure against the skin, the ground beneath a foot, or the position of a hand in darkness. This places it near the reactive end of the gradient introduced in the unit overview. Much of its machinery is built for control that cannot wait for deliberation, and some of its most consequential processing is therefore performed in the spinal cord rather than the cerebral cortex.
The shared sensory architecture developed in the overview is assumed here: physical energy is transduced at receptors, information is carried through ordered neuronal populations, the ascending pathways cross the midline (albeit at different levels), and thalamic relays project into mapped cortical fields embedded in recurrent networks. The task of this chapter is to specify how the body senses instantiate that plan—and to examine one place where the familiar textbook account has come under serious challenge.
This chapter concentrates on touch and proprioception. Pain and temperature are also part of somatosensation, but they will receive their own treatment in the subsequent Chapter 23. This is partly to keep this chapter at a reasonable length. But there are other reasons to delay a full accounting of pain. Pain is not simply ‘harder touch.’ It is an aversive valuation often associated with tissue damage that commands action. However, the peripheral and spinal connections for all submodalities of somatosensation, including pain, are similar and overlapping, and so will be introduced here in the current chapter.
Three senses under one name
Charles Sherrington divided the body’s sensory receptors into three broad functional classes in The Integrative Action of the Nervous System [@sherrington1906integrative]:
- Exteroception — sensing the external world through the body surface: touch, pressure, and the texture of objects against the skin.
- Proprioception — sensing the position and movement of the body itself: limb position, posture, and the state of muscles and joints.
- Interoception — sensing the internal organs and tissues, much of it outside awareness.
The classification places proprioception between the external world and the internal organs. Touch describes what is happening at the body surface; proprioception describes the body that is meeting that surface; interoception describes the body’s internal condition. These are genuine functional differences, but they are not three anatomically insulated systems.
Cutaneous and proprioceptive afferents from the trunk and limbs generally have their cell bodies in the dorsal root ganglia and enter the spinal cord through the dorsal roots. Visceral information travels through partly overlapping spinal pathways and through cranial routes, especially the vagus nerve. Somatosensory information from the face enters through cranial nerves, with first-order cell bodies in cranial-nerve ganglia rather than dorsal root ganglia. The peripheral machinery therefore overlaps without being identical.
The distinction between interoception and exteroception remains biologically useful. Interoceptive signals principally support regulation of the internal milieu; exteroceptive signals principally describe events beyond or at the surface of the body. Somatosensation reveals the seam between them because similar receptors, fibers, and physical variables can serve either domain depending on where they arise, where they project, and what control problem they help solve. A mechanically gated channel can report the texture of an object at the fingertip, the stretch of the bladder wall, or pressure within a blood vessel. The molecular operation is similar, but the signal enters a different regulatory loop.
The categories are therefore best understood by what the signals are used for, not by imagining three wholly separate sets of nerves. The interoceptive half of bodily sensation was developed in Unit II, where visceral and baroreceptor traffic reached the brainstem and hypothalamus to support homeostatic control. The present chapter follows the more outward-facing pair: touch at the skin and proprioception from muscles, tendons, and joints.
Exteroception and its receptors
Different submodalities of body sensation are transduced by different receptor endings. The classical scheme sorts the tactile mechanoreceptors of the skin along two axes: how rapidly they adapt to sustained stimulation and how deeply they lie in the skin.
| Rapidly adapting |
Meissner corpuscle (RA1) — flutter and light moving touch |
Pacinian corpuscle (RA2) — high-frequency vibration |
| Slowly adapting |
Merkel cell–neurite complex (SA1) — fine form, edges, and sustained pressure |
Ruffini ending (SA2) — skin stretch |
The logic of the table matters more than the four names. A rapidly adapting receptor emphasizes change: the onset and offset of contact, vibration, or movement across the skin. It becomes relatively quiet during steady pressure, which is one reason clothing soon fades from awareness. A slowly adapting receptor continues to respond during sustained deformation and therefore carries information about ongoing form and pressure.
Depth is related to spatial scale. Superficial endings generally have smaller receptive fields and support finer spatial detail. Meissner and Merkel receptors are especially dense in glabrous skin such as the fingertips, where small changes in contact matter for object manipulation. Deeper endings have larger receptive fields. Pacinian corpuscles are exquisitely sensitive to vibration that may propagate through a broad region of tissue, while Ruffini endings respond to skin stretch that accompanies hand shape and joint movement. Together the four classes sample different kinds of mechanical events rather than providing four redundant versions of touch.
A receptive field is the region of skin over which stimulation changes the activity of a sensory neuron. Receptive-field size and receptor density explain two-point discrimination. Two nearby contacts on a fingertip are likely to activate different afferents with small receptive fields and are therefore preserved as two spatial events. The same contacts on the back may fall within the receptive field of a single afferent and become indistinguishable. This is not merely a vague difference in sensitivity; it is a difference in the spatial sampling performed at the body surface.
Once two stimuli have been collapsed into the same peripheral signal, downstream processing cannot reconstruct the missing distinction. Cortical computation can sharpen or combine information that reaches it, but it cannot recover spatial detail that the receptor sheet never encoded. Spatial acuity is therefore greatest where receptive fields are small and afferents are densely packed. Those same regions receive disproportionate cortical territory. Fingertips, lips, and tongue gain an advantage at each stage: dense peripheral sampling, small receptive fields, and enlarged representation in the cortical map.
The four cutaneous classes all report events at the body surface—the exteroceptive face of somatosensation. But the same mechanical machinery is deployed deeper in the body, where identical physics answers a different question. A receptor that transduces deformation can report the texture of an object at the fingertip, the length of a contracting muscle, or the pressure inside a blood vessel; what differs is not the transduction so much as the control loop the signal enters. The remaining receptor systems are best grouped not as a miscellany of hardware but by that distinction of use.
Proprioception: sampling the body’s own configuration.
The muscle spindle is not a bare nerve ending buried in muscle but a small encapsulated organ. It contains specialized intrafusal muscle fibers, and the sensory afferent coils around their central region, which is itself non-contractile. Stretching the muscle stretches this central region, deforms the ending, and initiates the afferent signal; the spindle therefore reports muscle length and, dynamically, the rate at which length is changing. The Golgi tendon organ samples the complementary variable. Positioned in series at the junction of muscle and tendon, it reports muscle tension rather than length. The two measurements are genuinely independent—a muscle can be long and slack or short and straining—which is why proprioception needs both. This is the same principle already met at the skin: different receptors sampling different physical quantities, not redundant copies of one.
One feature of the spindle repays closer attention, because it reveals what proprioception is for. The central region of the intrafusal fiber is non-contractile, but its poles are contractile and are driven by their own dedicated motor supply, the gamma motor neurons, separately from the alpha motor neurons that drive the force-producing muscle around them. This separation solves a problem that would otherwise cripple the sense. As a muscle shortens, its spindles would go slack and fall silent—blind precisely when the limb is moving fastest. By contracting the poles of the intrafusal fiber, the nervous system re-tensions the spindle to keep it sensitive across the muscle’s working range. The receptor is thus not a passive gauge read out by the brain; it is continuously set by the brain, its sensitivity adjusted to the movement in progress.
That the spindle is set rather than merely read is the first sign of something this chapter will meet repeatedly: proprioceptive afferents are not reporters delivering limb position to awareness but sensors inside the loops that control movement, and the gamma system tunes them as the motor command changes. The same afferent will reappear at two more levels of that control hierarchy — carried to the cerebellum by the spinocerebellar tracts for online regulation, and forming the afferent limb of the stretch reflex, the shortest loop of all, which closes without the brain at all. Both are taken up later in this chapter. The full feedback architecture is the business of Unit 5, where the spindle and its gamma control return as working parts of the motor system rather than as entries in a receptor catalogue.
Interoception and nociception: the same transduction, other loops.
Two further receptor populations belong to somatosensation’s inward-facing side and are largely free (bare) nerve endings rather than encapsulated structures. Visceral stretch and pressure receptors—in the walls of hollow organs and blood vessels—feed the homeostatic control of the internal milieu; this interoceptive traffic was developed in the preceding unit, where it reached the brainstem and hypothalamus. Free nerve endings responsive to temperature and to tissue-damaging stimuli form the peripheral front end of pain, taken up on its own terms in Chapter 23. Here the seam between the sensory domains is at its most visible: a mechanically gated ending in a vessel wall and one at the fingertip may rely on similar channels yet serve regulation rather than perception. What sorts a receptor into one Sherringtonian class or another is not, in the end, the transducer—it is where the signal comes from and which control problem it is wired to solve.
Which afferents are fastest?
Somatosensory afferents differ greatly in conduction speed. Larger axons with thicker myelin conduct more rapidly. By convention, the principal classes are ordered Aα, Aβ, Aδ, and then unmyelinated C fibers. The largest myelinated fibers conduct at tens of meters per second, whereas many C fibers conduct at roughly a meter per second.
The ordering of the signals may appear counterintuitive. The fastest Aα fibers carry proprioceptive information from muscle spindles and tendon organs. Discriminative touch travels mainly on Aβ fibers. Nociceptive signals use the slower lanes: sharp, relatively rapid “first pain” travels on thinly myelinated Aδ fibers, while duller, lingering “second pain” travels on C fibers.
Subjective urgency might suggest that pain should receive the fastest wiring. Two features of the system explain why it does not. First, withdrawal can begin through short spinal circuits before nociceptive activity reaches cerebral cortex. The conscious experience of pain is not the first event in the protective response. Second, movement requires continuously updated estimates of limb position. A delayed estimate of muscle length or joint position would destabilize the control of a limb already in motion.
The body therefore spends its fastest wiring where moment-to-moment motor control needs it most, not where the resulting sensation feels most urgent. This ordering also anticipates a recurring principle of the movement unit: sensation is not merely information delivered to awareness. Much of it exists to close control loops.
Molecules that transduce force and temperature
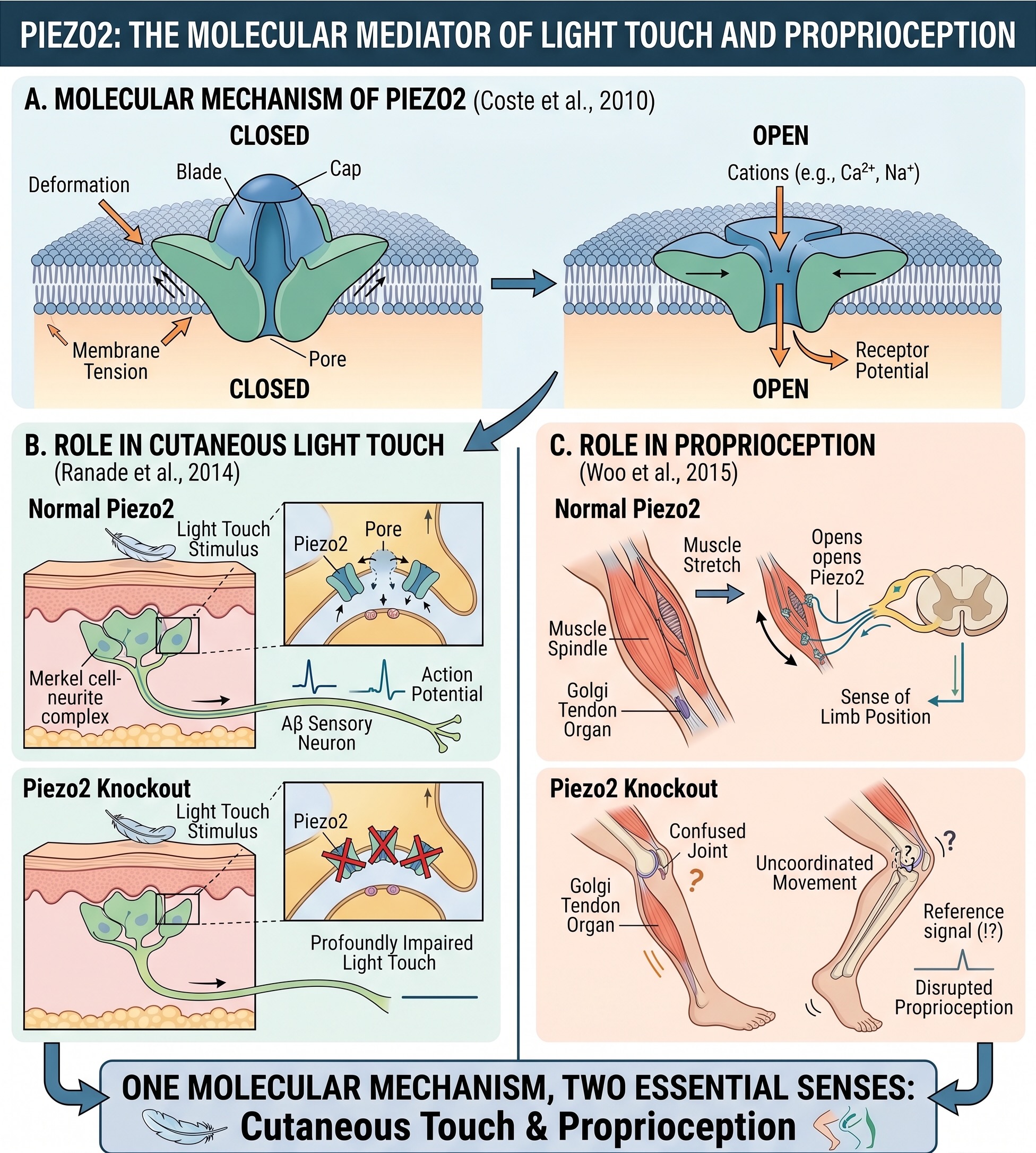
For much of the history of sensory physiology, a mechanoreceptor could be described only functionally: deformation somehow opened an ion channel and produced a receptor potential. The principal molecular force sensor is now known to be Piezo2, a membrane channel that opens when mechanical force deforms its local environment [@coste2010piezo]. Removing Piezo2 profoundly impairs light touch [@ranade2014piezo2touch], and removing it from proprioceptive neurons disrupts the sense of limb position [@woo2015piezo2prop]. The same molecular mechanism therefore contributes to both cutaneous touch and proprioception.
Temperature is transduced by members of the TRP channel family. TRPV1 responds to noxious heat and to capsaicin, which is why chili peppers produce a burning sensation. TRPM8 responds to cooling and to menthol, which is why mint feels cool [@caterina1997trpv1; @mckemy2002trpm8]. Other channels contribute across the thermal range, but these two provide particularly clear examples of a receptor protein linking a physical condition to an electrical response.
The 2021 Nobel Prize in Physiology or Medicine recognized David Julius and Ardem Patapoutian for discoveries of receptors for temperature and touch. The names worth retaining here are Piezo2, TRPV1, and TRPM8. The larger conceptual change is more important: the front end of touch and temperature is no longer a functional black box. Identified proteins now connect the physics of force and heat to the electrical language of the nervous system.
Discriminative touch is carried mainly by fast, myelinated Aβ fibers, but human hairy skin also contains C-tactile (CT) afferents: unmyelinated, slowly conducting low-threshold mechanoreceptors identified through microneurographic recordings from single peripheral fibers [@vallbo1999unmyelinated; @olausson2010ctreview]. CT afferents respond especially well to gentle, slowly moving touch near skin temperature—the physical profile of a caress. They are not prominent in the glabrous skin of the palms, where precise object manipulation dominates.
These fibers carry little precise information about form or location. People who lack normal fast tactile input but retain C-fiber input may report a diffuse sensation without accurate localization. CT afferents have instead been associated with the pleasant and social dimensions of touch and with activity in cortical regions involved in bodily feeling, including the insula.
Two cautions matter. First, the association between CT activity and pleasant touch should not be converted into a one-channel theory of affection. Pleasantness also depends on fast tactile signals, context, expectation, relationship, and the broader state of the organism, and the exact contribution of CT afferents remains under study [@schirmer2023ct]. Second, a pathway associated with the insula is not thereby purely “interoceptive.” The signal still begins with an event at the skin.
The CT system nevertheless complicates a simple account of touch as rapid spatial analysis. Somatosensation contains both a fast channel suited to discrimination and motor control and a slow channel linked to social and affective significance. The body senses again cross the boundary between exteroceptive description and interoceptive feeling.
The spinal cord: the body’s first sensory processor
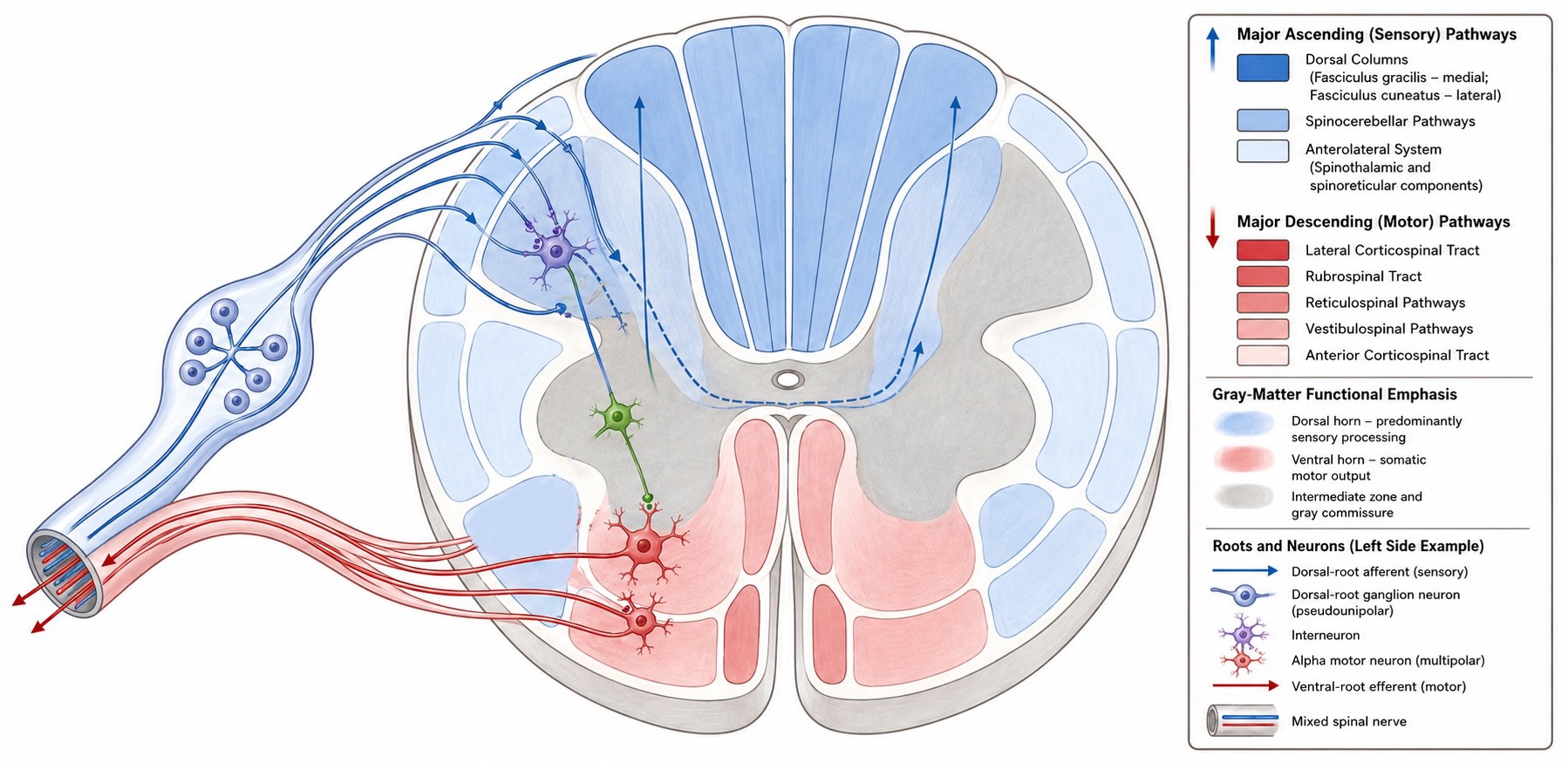
For the body rather than the face, sensory axons enter the central nervous system through the dorsal roots of the spinal cord, and their cell bodies lie in the dorsal root ganglia. The mnemonic AMPS captures the principal functional split: Anterior Motor, Posterior Sensory. Ventral roots carry motor axons out of the spinal cord; dorsal roots carry sensory axons in. The two join beyond the ganglion to form a mixed spinal nerve.
This sensory–motor division also preserves the developmental organization of the neural tube. The alar plate gives rise principally to sensory regions, while the basal plate gives rise principally to motor regions. The adult spinal cord therefore retains a visible trace of its embryological construction.
The first-order sensory neuron has an unusual shape. It is pseudounipolar: the soma sits off to the side in the dorsal root ganglion, while a single process divides into a peripheral branch and a central branch. In a typical central neuron, synaptic inputs arrive on dendrites and are integrated near the cell body. A somatosensory receptor neuron occupies a different position in the chain. There is no presynaptic neuron between its peripheral ending and the physical world.
The receptor ending must therefore convert pressure, stretch, temperature, or tissue damage into the neuron’s electrical response. The action potential is initiated at or near the peripheral ending and passes the offset soma on its way into the spinal cord. The first spike in the somatosensory chain can be born in a fingertip rather than in the spinal cord. Transduction and spike initiation occur at the body’s edge.
Three ascending pathways, different crossings and destinations
The most clinically consequential piece of anatomical bookkeeping is not simply that somatosensory pathways cross the midline, but where they cross and where they are going. Discriminative touch and conscious proprioception ascend toward cortex through the dorsal column–medial lemniscus system. Pain and temperature ascend mainly through the anterolateral system. Proprioceptive signals also diverge into spinocerebellar pathways that provide the cerebellum with rapidly updated information about the state of the body.
Proprioception therefore has two major central destinations. One pathway contributes to the conscious representation of body position in somatosensory cortex; another supplies the cerebellum with information used for the online regulation of movement.
The anterolateral system
The anterolateral system, including the spinothalamic tract, carries pain and temperature, together with some less discriminative forms of touch. The cell body of the first-order neuron again lies in the dorsal root ganglion, but its central axon follows a different course after entering the spinal cord.
Many pain- and temperature-sensitive afferents travel a short distance upward or downward in the dorsolateral tract of Lissauer before synapsing in the dorsal horn. The second-order axon then crosses the midline through the anterior white commissure, usually within one or a few spinal segments of the afferent’s entry. It subsequently ascends contralaterally in the anterolateral portion of the spinal cord.
The spinothalamic component reaches the thalamus and contributes to cortical representations of the location and intensity of a stimulus. Other components of the anterolateral system terminate in brainstem and midbrain structures involved in arousal, orienting, defensive responses, and the regulation of pain.
The defining sequence is therefore:
synapse in the spinal cord → decussation in the spinal cord → contralateral ascent toward the brainstem and thalamus
Both ascending cortical systems preserve body order, but they arrange it differently. In the dorsal columns, lower-body fibers lie medially and upper-body fibers are added laterally. In the anterolateral tract, fibers entering at progressively higher spinal levels are added more medially, leaving lower-body fibers relatively lateral. Somatotopic organization continues through the brainstem and thalamus and contributes to the ordered body map in somatosensory cortex.
Proprioceptive pathways to cortex and cerebellum
Proprioception begins with receptors that report the state of muscles, tendons, and joints, especially muscle spindles and Golgi tendon organs. Their first-order axons enter the spinal cord through the dorsal roots, but the resulting information is distributed to more than one central destination.
Some proprioceptive afferents send an ascending branch into the dorsal columns. This branch follows the dorsal column–medial lemniscus route: it ascends ipsilaterally to the medulla, crosses there, relays through the thalamus, and reaches somatosensory cortex. This pathway contributes to the conscious perception of limb position, movement, and the configuration of the body.
Other branches provide proprioceptive information to the cerebellum. For the lower trunk and lower limb, afferent collaterals synapse on neurons in Clarke’s column, a longitudinal group of neurons in the medial spinal gray matter. Axons from Clarke’s column enter the dorsal spinocerebellar tract, ascend largely without crossing, and enter the cerebellum through the inferior cerebellar peduncle. The cerebellum therefore receives information from the ipsilateral side of the body.
The corresponding route from the upper limb relays mainly through the accessory cuneate nucleus of the medulla and reaches the cerebellum through the cuneocerebellar tract. Other spinocerebellar pathways carry related information about the activity of spinal circuits and have more complicated crossing patterns, but the Clarke’s-column pathway illustrated here remains predominantly ipsilateral.
These cortical and cerebellar projections serve different purposes. The cortical route supports conscious judgments about where the body is and how it is moving. The cerebellar route supplies a rapid, continuously updated estimate of the state of the limbs and trunk that can be compared with ongoing motor commands. Much of this cerebellar processing contributes to movement without entering conscious awareness.
The clinical consequence of the crossings
Because these pathways cross at different levels, a lesion affecting one half of the spinal cord produces a characteristic dissociation. Discriminative touch, vibration, and conscious proprioception are impaired ipsilateral to the lesion below the damaged level because the dorsal-column fibers have not yet crossed. Pain and temperature are impaired contralaterally, usually beginning one or a few segments below the lesion, because the relevant second-order neurons have already crossed within the spinal cord.
Damage to the dorsal spinocerebellar tract can also impair coordination on the ipsilateral side. This deficit may appear as limb ataxia, although it can be difficult to distinguish from the effects of weakness or loss of conscious proprioception in a more extensive spinal injury.
The opposed loss of dorsal-column and anterolateral sensation is the characteristic sensory component of Brown-Séquard syndrome. A complete hemicord lesion also interrupts descending motor pathways, producing upper-motor-neuron weakness ipsilateral to the lesion below the damaged level. The syndrome makes the internal geometry of the spinal cord visible at the bedside: dorsal-column information crosses late in the medulla, pain and temperature cross early in the spinal cord, and the principal spinocerebellar route from Clarke’s column remains on the same side as the body it represents.
Reflexes: processing in the cord
Because somatosensation serves reactive control, some of its processing never reaches the brain. The stretch reflex provides the cleanest example. Afferents from a muscle spindle enter the spinal cord and make a direct excitatory connection onto alpha motor neurons supplying the same muscle. Stretch increases spindle firing; the motor neurons respond; the muscle contracts against the perturbation. No brain is required. The loop detects a deviation and begins correcting it within the spinal cord.
The circuit is more than a simple wire. Interneurons coordinate the response by inhibiting antagonist muscles and distributing activity across motor pools, so that even the most elementary reflex embodies an organized relationship between sensory input and motor output. What the stretch reflex lays bare is the control loop itself, stripped to its minimum: a sensor measures a variable, and its signal drives an action that opposes the measured deviation, all of it closing in the cord with no brain in the path. This is the primitive from which the rest of the book’s control systems are built. Whether such a loop is deployed reactively — correcting an error once it appears, as here — or predictively, anticipating the error before it arrives, is the distinction between homeostasis and allostasis drawn in the overview, and it is a distinction about how the loop is used, not about the loop itself.
If the stretch reflex is a loop with no controller, the cord’s central pattern generators (CPGs) are a step further still: circuits that generate their own reference. Under appropriate conditions, spinal circuitry isolated below a transection can produce coordinated stepping when the limbs meet a moving treadmill, the rhythm arising from the network’s own connectivity rather than from a descending command issued step by step. Sensory feedback shapes the timing and force of each cycle, but the pattern does not have to be specified from above. Here the overview’s claim about nervous tissue becomes concrete: the wiring does not implement an algorithm for walking that is stored somewhere and executed; the wiring is the walking. The rhythm is a property of the architecture.
The spinal cord is therefore not a cable connecting body to brain but a sensorimotor structure in its own right, holding both the loops that correct disturbances and the programs that generate movement. This chapter meets these circuits only as the local, fast end of somatosensation; they return as working machinery in the unit on movement. Their presence here follows directly from the character of the system this chapter has been describing — a set of body senses built, before anything else, for action that cannot wait.
From thalamus to somatosensory cortex
The thalamic relay
Body sensation relays principally through the ventral posterior thalamus:
- VPL (ventral posterior lateral) receives information from the body through the medial lemniscus and anterolateral pathways.
- VPM (ventral posterior medial) receives information from the face, principally through trigeminal pathways.
The thalamic relay preserves somatotopic order and projects onward to primary somatosensory cortex. As emphasized in the unit overview, this is not a passive one-way transfer. Corticothalamic feedback and local thalamic inhibition make the relay part of a recurrent circuit. The cortex receives a mapped input from the thalamus while simultaneously helping regulate what the thalamus transmits.
Primary somatosensory cortex: four maps, not one
Primary somatosensory cortex, or S1, occupies the postcentral gyrus. It is not a single uniform map but a set of adjacent fields—Brodmann areas 3a, 3b, 1, and 2—that emphasize different inputs and computations:
- Area 3a emphasizes proprioceptive input from muscles and joints.
- Area 3b is the principal cortical recipient of cutaneous input.
- Area 1 receives strongly from area 3b and is especially responsive to texture and movement across the skin.
- Area 2 combines cutaneous and proprioceptive information important for representing object size and shape.
The sequence is partly hierarchical but strongly recurrent. Area 3b supplies important input to areas 1 and 2, while connections within S1 and feedback from later regions continually reshape the activity of each field. Even within “primary” cortex, the body is represented several times and in several forms.
Vernon Mountcastle’s single-unit recordings in cat somatosensory cortex revealed another important feature of S1. Neurons encountered along a vertical electrode penetration tended to respond to the same class of peripheral receptor and nearly the same receptive-field location, whereas oblique penetrations crossed abrupt changes in modality or body location [@mountcastle1957columnar]. This observation helped establish the cortical column as a recurring principle of cortical organization. Hubel and Wiesel later described related columnar organization in visual cortex [@hubel1962receptive]. A general principle of cortical architecture was first seen especially clearly in the body senses.
Figure 4.1.3. Vernon B. Mountcastle (1918–2015), whose single-unit recordings revealed the vertical, columnar organization of primary somatosensory cortex. [Portrait to source.]
Directly across the central sulcus lies primary motor cortex, which carries its own ordered representation of the body. Sensory and motor representations of the same body part therefore occupy neighboring cortical territory. The arrangement places information about the hand beside circuitry that helps control the hand, matching the action-centered purpose of the system.
The somatosensory homunculus
The ordered representation of the body across S1 is commonly depicted as the sensory homunculus. The resulting “little human” is grossly distorted: lips and hands occupy large expanses of cortex, while the trunk and proximal limbs occupy much less. Cortical territory reflects receptor density, spatial acuity, and behavioral use rather than the physical size of the body part.
The familiar map emerged from a long history of stimulation and lesion studies, including work by Fritsch and Hitzig, David Ferrier, and many others. Wilder Penfield’s stimulation of the exposed cortex in awake patients undergoing epilepsy surgery gave the homunculus its familiar modern form [@penfield1950cerebral]. When stimulation evoked a sensation in the hand, face, or leg, the surgical team could relate that report to a location on the cortical surface.
The drawing should not be mistaken for a sharply bordered miniature body. Representations overlap, vary across individuals, and depend on how they are measured. Different S1 fields also contain partly different maps, as the four-area organization makes clear. The large-scale somatotopic order is nevertheless robust: leg and foot are represented medially, the trunk and arm more dorsolaterally, and the face most laterally.
Secondary somatosensory cortex
The secondary somatosensory cortex (S2) lies in the parietal operculum along the lateral sulcus. It receives input from S1 and from the thalamus. Whereas S1 is dominated by the contralateral body, S2 contains a more bilateral representation, supported in part by callosal communication between the two hemispheres.
S2 neurons often have larger receptive fields and can combine information across several digits or both sides of the body. The strict contralateral organization of the earliest cortical map therefore softens as later regions integrate information over larger portions of the body and across longer sequences of touch. This is an early step toward recognizing an object by handling it rather than merely registering contact at a point on the skin.
The cerebellar route
The spinocerebellar tracts carry muscle-spindle and tendon-organ information to the cerebellum for the online regulation of movement. Functionally, this information remains predominantly ipsilateral: the cerebellum receives information about the same side of the body that it helps control. Some spinocerebellar fibers remain uncrossed, while others cross twice and thereby return to the original side.
Their same-sided organization contrasts with the prominent contralateral arrangement of conscious sensory and motor pathways. The contrast makes an obvious question unavoidable: why did vertebrate sensory and motor systems adopt crossed pathways in the first place?
The contralateral organization of many vertebrate sensory and motor pathways is anatomically secure, but its evolutionary and developmental explanation remains unsettled. A same-sided arrangement can appear simpler, yet many pathways cross while others—including much of the olfactory system and the functionally ipsilateral spinocerebellar system—do not.
Functional accounts propose that crossing improves the alignment of sensory maps with the limbs or muscles that act on the represented space. A visual event in one half of the world is represented in the opposite hemisphere, where motor commands to the contralateral limbs may be organized in a compatible spatial register. Such accounts offer a possible selective advantage, but they do not readily explain the detailed pattern of crossed, uncrossed, and double-crossed pathways.
Developmental and evolutionary accounts propose that the arrangement is a residue of an ancestral twist in the vertebrate body plan. In the axial-twist hypothesis, the anterior head is proposed to have rotated relative to the rest of the body, with later developmental compensation producing the observed crossings [@delussanet2012axial]. The related somatic-twist hypothesis proposes a different sequence of rotation [@kinsbourne2013somatic]. These models attempt to explain the anatomy directly, including some exceptions, but the proposed twist and its genetic-developmental basis have not been established.
No account has gained general acceptance. Decussation is therefore a conspicuous example of an anatomical fact whose ultimate explanation remains open.
When sensation survives but recognition fails
The pathways traced so far deliver an ordered description of events at the skin: where contact occurred, how hard, how fast it moved, whether it is still there. But knowing that something narrow and cold with jagged teeth is lying across the fingers is not the same as knowing it is a key. Somewhere beyond the point where the body surface has been faithfully described, touch becomes recognition.
That transition can fail on its own. An agnosia is a loss of recognition that cannot be explained by a loss of sensation. The receptors work, the pathways deliver, the elementary percept arrives intact—and the object still is not identified. The word comes from the Greek gnosis, knowledge, with the negating a-: literally, not knowing. Agnosias are among the most informative disorders in neuropsychology, because they separate two things that ordinary experience presents as one. Recognizing your keys in a dark pocket feels like a single act. A patient in whom sensation survives while recognition fails shows that it is not.
Tactile agnosia is the failure to recognize objects by touch despite intact elementary sensation. The qualification carries the entire weight of the term. A patient who cannot identify a key because dorsal column damage has degraded the tactile signal has no recognition disorder at all; the information never arrived. This is the same constraint met earlier at the receptor sheet, where two contacts collapsed into one afferent could not be recovered downstream. Before any claim about higher-order processing can be made, elementary sensation must be shown to be normal—which is why the clinical rule is that cortical sensory testing means nothing until light touch, pinprick, temperature, vibration, and proprioception have been tested first and found intact. (Clinical sources also use the older term astereognosis, sometimes for this disorder and sometimes for any failure of tactile recognition whatever its cause. The distinction that matters is the one above.)
The case of E.C.
The clearest demonstration comes from a patient studied in detail by Reed, Caselli, and Farah [@reed1996tactile; @reed1994nature]. E.C. sustained a small, well-localized infarct of the left inferior parietal cortex. Afterward she could no longer identify objects held in her right hand. Asked to name forty common items by touch, she managed roughly half with the affected hand and nearly all of them with her left.
Her errors were revealing. She identified a pine cone as a brush and a snail shell as a bottle cap—structural confusions, the responses of someone building an incorrect shape rather than someone feeling nothing.
Elementary sensation was normal in both hands: light touch, pinprick, temperature, proprioception, two-point discrimination, and vibration, the last confirmed with instrumented threshold testing rather than bedside examination alone. Intellect, language, and motor function were unaffected.
The unilateral deficit is what makes the case decisive. E.C.’s unimpaired hand serves as a control within the same patient, on the same day, with the same brain. No appeal to general intelligence, motivation, or task difficulty can explain why one hand recognizes a key and the other does not.
The investigators then worked to eliminate the deflationary explanations rather than assuming their absence. It had been argued that tactile recognition failures are really failures of general spatial ability, since touch must assemble a shape from a sequence of contacts [@semmes1965nontactual]. E.C. was given the same map-following task used to support that claim and scored above the average of the original control group. She also performed normally on visual tests of assembling objects from separately viewed parts.
Three further results narrowed the deficit:
- Shape, not location. Asked to locate a raised dot within a palpated square, E.C. performed normally with both hands. Asked to identify shapes, she failed with the agnosic hand alone—a dissociation within touch itself.
- Not faulty exploration. Her hand movements followed normal exploratory patterns, and guiding her finger around each contour did not rescue performance.
- Scaling with complexity. She estimated the length of a dowel as accurately as controls and judged which dimension of a polygon was longest without error, but fell to chance on complex unfamiliar shapes. The impairment grew as the shape demanded more.
What remains after these eliminations is a deficit specific to tactile shape perception: not sensation, not spatial cognition, not exploration, not object knowledge. E.C. could still draw an object she had failed to name, and her drawings showed the shape she had actually constructed—outlines roughly right, internal details missing or duplicated.
What the case establishes
Between the arrival of tactile signals and the recognition of an object lies a stage of processing that belongs to touch specifically and can be lost on its own. The homunculus and the ascending pathways describe how the body surface is represented. They do not, by themselves, explain how a handful of sequential contacts becomes a key. E.C. shows that the further step exists, is anatomically vulnerable, and depends on parietal cortex beyond the primary map.
This is the first agnosia in this book, and it is worth registering the shape of the argument, because it recurs. A single well-characterized patient is not weak evidence when the dissociation is sharp and the alternatives have been tested rather than assumed. Recognition disorders in vision are more dramatic and better known, and ?sec-visual-agnosia takes them up in detail. The logic that makes them interpretable is the logic set out here.
How fixed is the map?
The somatosensory homunculus invites a natural inference: a body map this orderly must be permanently hard-wired. A long tradition of research instead described adult cortical maps as highly plastic, and for several decades that account—deprivation frees cortical territory, neighboring inputs expand into it, and phantom sensations reveal the perceptual consequence—became the standard demonstration that adult cortex reorganizes dramatically after injury. More recent work, using stronger designs, has constrained that account substantially. The disagreement is not simply whether cortex can change, but what kind of change should count as a reorganized map—and, as we will see, the two bodies of evidence are not equally weighty.
What the disagreement actually concerns
The revised evidence strongly challenges the strongest “takeover” account, but it does not establish that adult cortex is static. Much of the confusion comes from grouping genuinely different phenomena under the single word reorganization:
- increased influence of inputs that were already present but previously weak;
- altered gain or receptive-field size within an existing circuit;
- changes in how downstream regions read a preserved representation;
- structural rewiring that supplies genuinely new input;
- developmental changes that occur while cortical areas are still being specified.
These processes need not rise and fall together. A neuron may respond more strongly to a neighboring body part because a previously weak input has been unmasked, not because the neuron’s original circuitry has been erased and rebuilt. A preserved hand representation may also be read differently by later cortical regions once the hand is gone. Both outcomes are forms of plasticity, but neither requires wholesale reassignment of cortical identity.
The distinction that matters most is between the developing brain and the adult one. Developmental input-dependence is real and dramatic: in a classic transplantation experiment, late-embryonic visual cortex placed into neonatal somatosensory cortex developed barrel-like units characteristic of rodent whisker cortex [@schlaggar1991potential]. Cortical identity is therefore not specified entirely in advance. But that a developing area can be shaped by its inputs says little about whether an adult map can be reassigned to a new body part on the same scale. Digit amputation, complete deafferentation, congenital absence of a limb, temporary disuse, and adult surgical amputation are not interchangeable interventions, and evidence from one does not transfer freely to another.
The most defensible current conclusion is therefore more constrained than either extreme. Adult somatosensory responses can change, sometimes substantially, but the original body representation may persist beneath altered response strength, access, or readout. How much adult map change reflects unmasking, gain adjustment, downstream interpretation, or genuine rewiring remains an active question [@sparling2024bridging].
This is a case in which stronger evidence makes the textbook account less tidy. That is not a failure of the field; it is what happens when a compelling explanatory story is separated into several testable claims and those claims are examined with better designs.
The classical and revised pictures are often presented as two sides of an open debate. They are better understood as evidence of unequal weight, which is why the balance of opinion has shifted.
The evidence that built the plasticity account came largely from animal recordings and cross-sectional human comparisons:
- Digit amputation in adult monkeys. After a finger was amputated, Merzenich and colleagues found that cortical responses driven by neighboring fingers expanded into territory formerly associated with the missing digit [@merzenich1984digit]. The deprived territory did not simply fall silent.
- Long-term deafferentation. Pons, Kaas, and colleagues reported that, years after an arm had been deafferented, stimulation of the face drove neurons in a large region formerly associated with the hand and arm [@pons1991massive]. Because face and hand are neighbors in the map, this appeared to show large-scale invasion of deprived territory.
- Referred sensations in phantom limbs. Ramachandran reported that touching the face could evoke sensations referred to a phantom hand after amputation [@ramachandran1998phantom], fitting the proposal that facial input had gained access to the neighboring hand representation.
The evidence that constrained it came from asking what those measured changes actually demonstrate [@makin2017stability; @makin2023against]:
- Attempted movements of phantom fingers can evoke organized activity resembling the intact hand representation, implying the missing hand’s functional organization persists.
- Stimulation of residual nerves or of cortex can evoke sensations assigned to the missing hand, again implying the representation remains accessible.
- Neighboring body parts already produce weak, overlapping activity outside the territory they dominate. Once the missing hand can no longer be stimulated, a “winner-takes-all” readout can make those pre-existing inputs look like newly invaded territory without any circuit having been rebuilt.
- Most foundational human studies were cross-sectional, comparing amputees with controls. Such designs cannot reveal whether a difference was caused by amputation or was present beforehand.
The decisive test was longitudinal. Schone, Makin, and colleagues followed three adults before a planned arm amputation and for as long as five years afterward, comparing activity for the hand before surgery, the phantom hand after, and the lips throughout. They found stable hand and lip representations in primary sensorimotor cortex and no sign that the lips had spread into the missing-hand territory [@makin2025stable]. The sample was necessarily small, but the design addressed the central weakness of every earlier human comparison: the same brains were measured before and after the loss. A single well-designed prospective study of this kind outweighs a larger number of cross-sectional ones, and it is this asymmetry—not a mere shift in fashion—that has moved the field.
The body you feel is a model, and you can fool it
Bodily experience is constructed from multiple streams of evidence rather than read directly from a single sensory map. Several body illusions make that construction visible.
In the rubber hand illusion, a participant’s real hand is hidden while a visible artificial hand is placed in a plausible position. When the real and artificial hands are stroked synchronously, many participants begin to experience the seen hand as part of their body. The perceived position of the hidden hand may also drift toward the artificial one. Vision, touch, and proprioception have been combined into a body estimate that differs from the physical arrangement.
The effect is not simply a verbal judgment that the rubber hand “looks like mine.” Threatening the artificial hand can evoke defensive reactions, indicating that the temporary body model has consequences for action. The nervous system resolves a conflict among sensory signals by adopting a plausible common cause: the seen hand is being touched when and where touch is felt, so the hand is treated as belonging to the body.
The Pinocchio illusion turns proprioception against touch. Vibration of the biceps tendon can make muscle-spindle signals indicate that the forearm is extending. When a blindfolded participant simultaneously touches the nose, the nervous system must reconcile a finger that remains in contact with a hand that seems to be moving away. The resulting percept can be an elongating nose—an impossible body that makes the sensory signals mutually consistent.
The cutaneous rabbit reveals the importance of temporal and spatial inference within touch itself. Rapid taps delivered first near the wrist and then near the elbow can be felt as a sequence hopping through untouched positions between them. Activity in somatotopically intermediate regions of S1 can accompany the illusory taps. The sensory system is not merely registering each contact independently; it is inferring the most plausible trajectory across the skin.
Together, these effects show that the experienced body emerges from interactions among cutaneous, proprioceptive, visual, and temporal signals in a distributed network that includes S1 but extends well beyond it. They do not by themselves show that the anatomical map of S1 is structurally redrawn within minutes. A constructed body representation and a rapidly rewired primary map are different claims.
The same constructive capacity is important for neuroprosthetics. When visual, motor, and artificial sensory feedback are brought into register, a prosthetic device can become incorporated into the user’s body representation, improving the naturalness and control of the device. Disorders such as tactile agnosia reveal the complementary failure: sensation may remain intact while the object or body significance of that sensation is lost.
Looking ahead
Somatosensation occupies the reactive, body-contact end of this unit. Its receptors sample force, temperature, and the state of muscles and tendons; its fastest afferents serve motor control; its spinal circuits begin correcting disturbances before cortical perception is complete; and its ascending pathways preserve ordered representations through thalamus and cortex. The resulting body map is robust, but the mechanisms and limits of its adult plasticity remain under active debate.
The next chapter turns to pain. The peripheral fibers and ascending pathway have now been introduced, but pain adds something more than sensory description: aversiveness, valuation, and an urgent demand for action. It therefore provides the first direct bridge from sensation toward the motivational and action systems developed later in the book.
Reasonably settled:
- Sherrington’s distinction among exteroceptive, proprioceptive, and interoceptive receptors remains a useful functional organizer, although the categories use partly overlapping peripheral machinery.
- Tactile mechanoreceptors differ in adaptation rate, receptive-field size, and depth. Piezo2 is a principal force transducer for touch and proprioception, while TRP channels contribute to temperature transduction.
- Conduction speed follows axon diameter and myelination. Proprioceptive Aα fibers are fastest, tactile Aβ fibers follow, and nociceptive Aδ and C fibers are slower.
- First-order somatosensory neurons from the body are pseudounipolar dorsal-root-ganglion cells whose action potentials begin near peripheral receptor endings.
- The dorsal column–medial lemniscus and anterolateral systems cross at different levels. Their dissociation produces the characteristic sensory pattern of Brown-Séquard syndrome.
- Body and face relay principally through VPL and VPM, respectively, before reaching the somatotopic fields of S1. S2 contains a more bilateral representation.
- The spinal cord performs consequential local processing through reflex circuits and central pattern generators.
Genuinely unsettled, and presented as such:
- How adult cortical maps change after injury. The strongest longitudinal evidence argues against wholesale takeover of the missing-hand representation after adult amputation, but altered gain, unmasking, receptive fields, access, and downstream readout can still produce substantial functional change.
- How much pleasant and social touch depends specifically on C-tactile afferents. Their contribution is real, but affective touch is not reducible to a single peripheral channel.
- Why so many vertebrate sensory and motor pathways decussate. Functional and evolutionary-developmental hypotheses exist, but none has gained general acceptance.
Most of the chapter’s anatomy—the receptors, peripheral neurons, tracts, crossings, thalamic relays, and large-scale cortical map—is well established. The principal open questions concern what the adult map can become and why the pathways were crossed in the first place.