48 A Murder of Brains
Niche Construction, Cultural Inheritance, and the Worlds That Build Brains
48.1 The loop closes
Imagine trying to explain a honey bee by removing one worker from the hive and studying it alone. You could describe its eyes, wings, antennae, gut, genome, and brain. You would learn a great deal about bees. You would also miss the fact that organizes nearly everything you had measured.
A honey bee is built to live in a colony.
The comb organizes brood, food, and movement. Workers collectively ventilate the nest, defend its entrance, gather and redistribute food, and hold the brood within a comparatively narrow thermal range. A larva’s developmental future depends not only on the genes inside its cells but on what other bees feed it and on the social conditions in which it develops. The hive is simultaneously a physical structure, a food store, a nursery, a social organization, and a regulatory system. Bees build it, and it helps build bees.
That reciprocal sentence contains the central idea of this final unit.
Chapter 2 asked why an organism should have a brain at all. The answer began with adaptive movement and widened into control. A brain regulates a body in a world: it allocates internal resources, selects among possible actions, learns from consequences, and prepares for disturbances that have not yet arrived. Homeostasis became allostasis—stability achieved through anticipatory change rather than by holding every variable fixed.
This unit adds the return path. Organisms do not merely encounter environments. They choose among them, alter them, and sometimes leave those alterations behind. A beaver changes a stream into a pond. Termites construct a mound that modifies gas exchange, moisture, and temperature. Birds build nests; spiders make webs; animals dig burrows, wear paths, store food, cultivate fungi, and transport seeds. Some changes vanish when the animal leaves. Others persist long enough to shape the development and survival of offspring that did not make them.
This feedback is called niche construction: organisms modify or stabilize features of their surroundings and thereby alter the developmental conditions and selection pressures experienced by themselves and other organisms [@laland2016; @laland2017]. The empirical claim is secure, although evolutionary biologists continue to disagree about how much theoretical novelty the term adds. Nothing here requires replacing natural selection. The narrower correction is enough: the environment that selects an organism is not always an independent backdrop. It may be partly the product of organisms that came before.
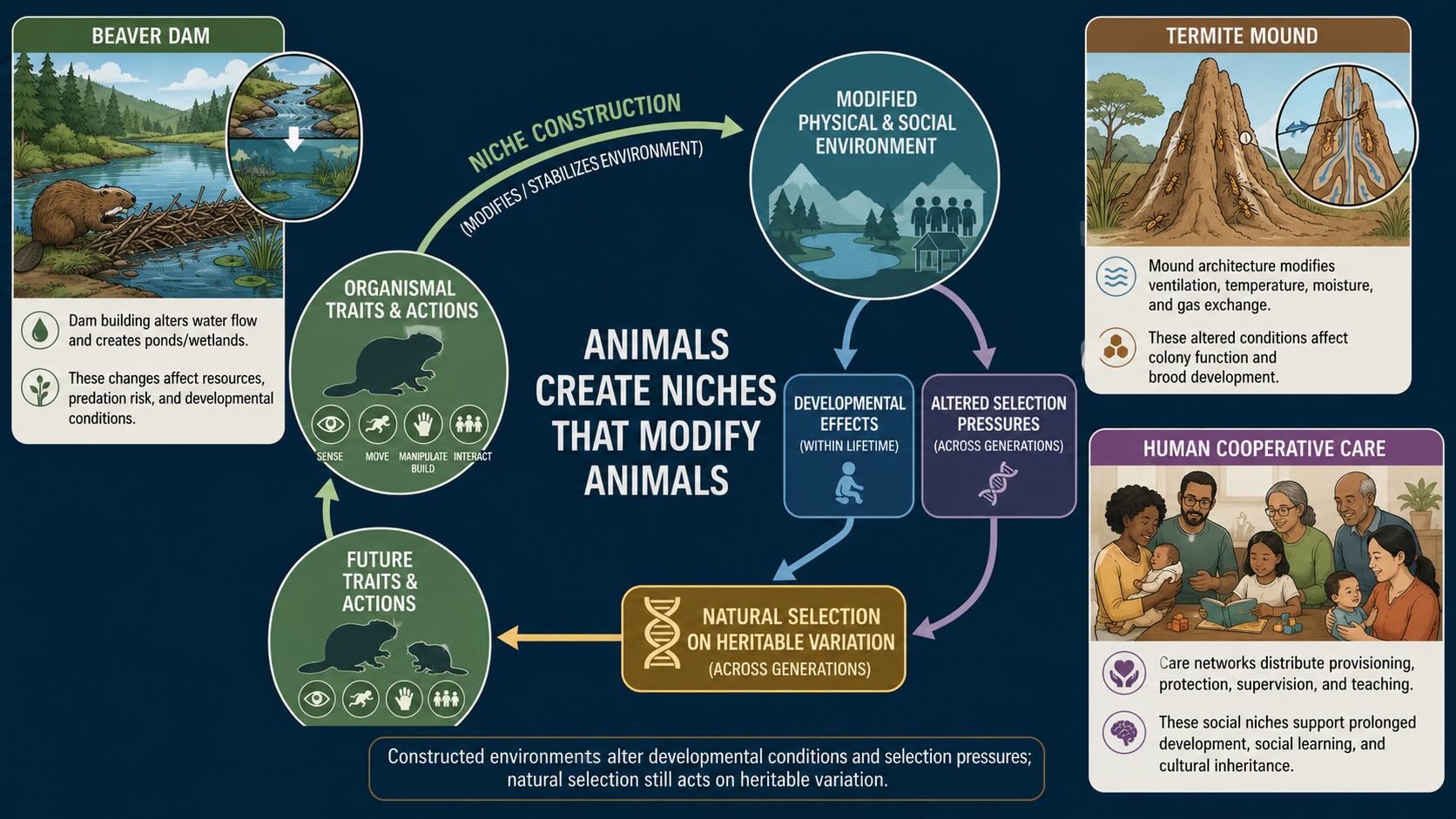
Animals create niches that modify animals.
Humans did not invent that loop. We intensified it, especially through a social problem introduced near the beginning of the book. Human offspring combine costly brain development with unusually prolonged dependence. No single universal childcare arrangement follows from that fact, and human mothers remain central across societies. Yet children are commonly provisioned, protected, carried, supervised, and taught by a wider network that may include fathers, grandparents, siblings, other relatives, and unrelated partners. Cooperative care is therefore best treated not as a tidy claim that two parents are literally incapable of raising a child, but as a flexible social infrastructure that repeatedly helped distribute the costs of human development [@hrdy2016; @burkart2025].
That infrastructure carries more than calories. It carries protection, labor, local knowledge, skills, and expectations. A social niche capable of pooling those resources can support a longer period of neural development and learning. A longer childhood, in turn, increases dependence on information accumulated by other people. Brains, bodies, social practices, and constructed environments therefore changed together. Culture arose from evolved organisms, and then became part of the environment within which later organisms developed and evolved.
The title of this unit is meant to shift the unit of analysis. A single brain remains indispensable, but many human capacities depend on a population of brains coupled by care, exchange, imitation, language, artifacts, and institutions. The relevant system is not a harmonious group mind. It is a distributed and often contested network in which resources and information can persist beyond any one individual. A flock of crows is called a murder. The phrase a murder of brains names the network on which a human life depends.
This overview follows the loop from physical niche construction through social learning, cultural inheritance, and gene–culture coevolution. Chapter 49 treats language not as an isolated faculty but as a biological and culturally inherited system for coordinating action and moving knowledge through a group. Chapter 50 turns to the most consequential objects in that group: other agents whose goals, knowledge, loyalties, and likely actions must be estimated.
Before reaching language or mind-reading, however, we should spend some time with animals that build.
48.2 Three builders
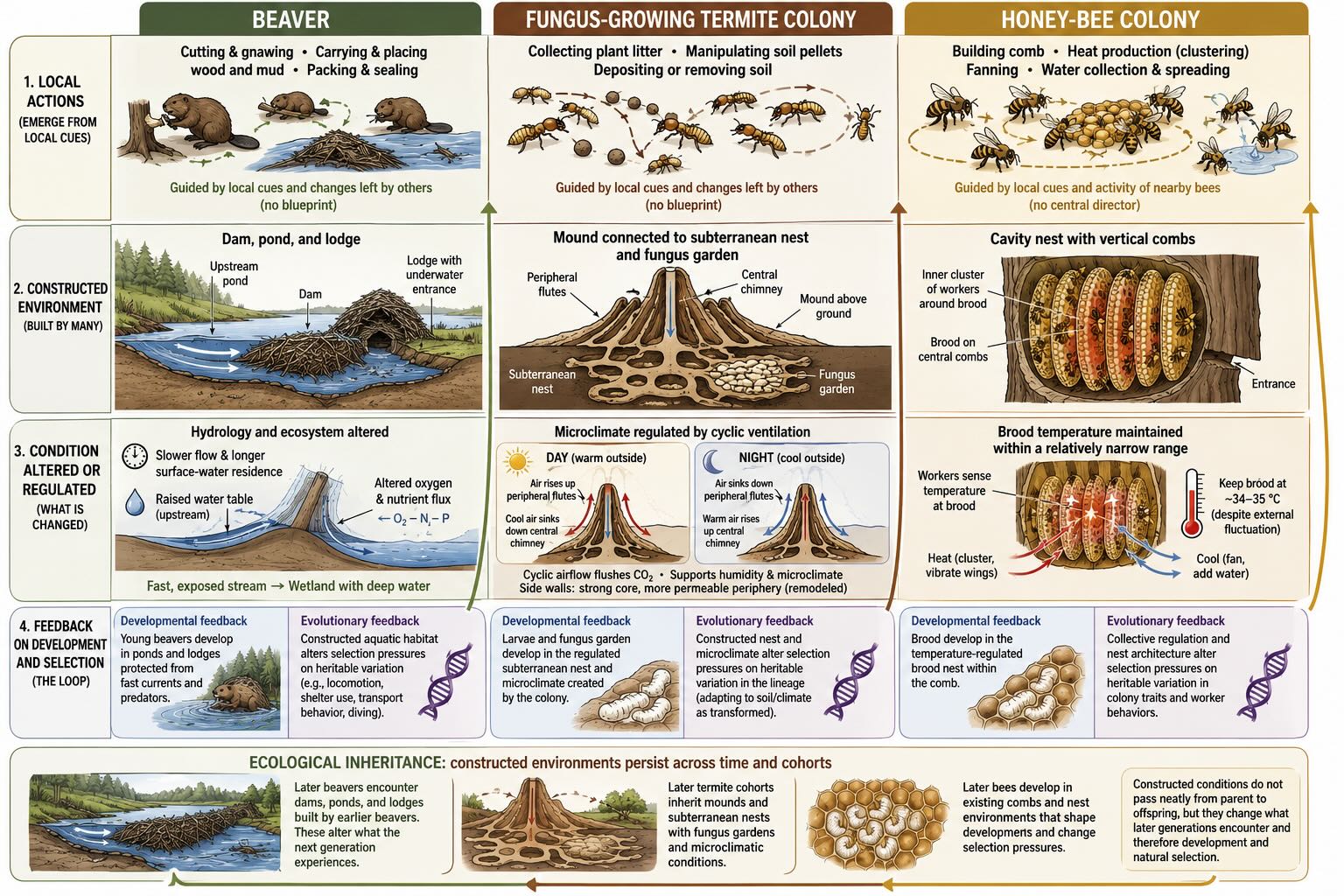
Organisms alter their worlds in countless ways, but a few cases make the loop unmistakable because the constructed object is large, durable, and obviously regulatory. Three are worth keeping together, because their convergence is the point: two are insects with brains smaller than a grain of rice, and one is a mammal, yet all three manufacture an environment that then feeds back on the manufacturer.
A beaver dam slows moving water, raises the water table, increases residence time, changes oxygen and nutrient flux, and converts a fast, exposed stream into a wetland with deep water around a protected lodge [@dewey2022]. The animal has not merely found a suitable habitat. It has built one. And the built habitat changes which beaver traits are valuable: lodges with underwater entrances, webbed hind feet, valves that close the ears and nostrils, and behaviors for transporting wood through water all make sense in a world partly constructed by beavers. We should resist turning this into an evolutionary history in which dam building caused each anatomical feature, because evolution is almost never that clean. The point is causal reciprocity. Beaver traits make dams possible, and dammed environments alter the value of beaver traits.
A termite mound performs similar work through architecture rather than hydrology. Fungus-growing termites cultivate a food source that requires a humid, chemically tolerable environment, and their mounds are not inert heaps of soil. The structure harnesses daily temperature oscillations to move air and flush carbon dioxide from the nest, while the mound walls balance structural strength against permeability [@king2015; @zachariah2020]. No termite possesses a blueprint of the completed structure. Local actions, guided by local cues and by changes left by other workers, generate a building that performs colony-level regulatory work. The mound then becomes part of the termites’ environment — sheltering the fungus garden, channeling air, storing moisture, and changing the conditions under which larvae develop. A termite lineage that depends on cultivated fungus is no longer adapting to soil and climate alone. It is adapting to soil and climate as transformed by termite construction.
A honey bee colony makes the same loop visible in the social dimension, and here the regulated variable is impossible to locate in any single animal. A colony can contain tens of thousands of workers, yet no worker directs the whole. Individuals respond to temperature, odor, vibration, food availability, the presence of brood, and the activity of nearby bees. From these local interactions emerge collective functions: foraging, defense, food allocation, nest repair, waste removal, and temperature control. When cold threatens, workers generate and conserve heat; when heat rises, they fan and distribute water for evaporative cooling, holding the brood nest comparatively stable despite large fluctuations outside. The wax comb adds passive insulation and imposes a physical organization on the social network.
Set side by side, the three builders teach one lesson with clarity. A mammal regulates body temperature through blood flow, sweating, shivering, posture, and behavior — all inside one skin. A bee colony regulates brood temperature by moving and activating many separate bodies inside a built enclosure. The regulated variable belongs to the colony’s nursery; the effectors are individual bees; the architecture couples them. The control circuit does not fit inside any single animal. That is the point this unit will make repeatedly: regulation that was once the work of one nervous system becomes the work of many bodies, artifacts, and inherited structures acting together.
This persistent alteration of the world has a name. Offspring inherit genes from their parents, but they may also inherit a modified environment — a nest, burrow, dam, depleted food patch, cultivated microbial community, altered soil, or changed population of prey and predators. This is ecological inheritance. It is not encoded in DNA, and it need not pass neatly from parent to offspring. Nevertheless, it changes what the next generation encounters, and therefore changes development and selection [@kendal2011]. An inherited environment need not be external to evolution. It can be one of evolution’s products.
Niche construction has generated an unusually heated argument, in part because two different questions are easily confused.
The first is empirical: do organisms alter environments in ways that change selection? The answer is yes. Dams, nests, agriculture, oxygen-producing photosynthesis, antibiotic use, and host modification of microbial communities all do this.
The second is theoretical: does this require a reformulation of evolutionary theory? Here opinions differ. Advocates of niche-construction theory argue that standard presentations assign causal priority to an external environment and treat organismal activity as merely an outcome of prior selection. They want reciprocal causation and ecological inheritance placed beside natural selection rather than hidden inside the category of “environment.” Critics reply that ordinary evolutionary theory has always been capable of modeling organisms that alter their surroundings, and that giving the phenomenon a new name does not create a new evolutionary mechanism [@scottphillips2014].
We do not need to settle that dispute, and you should know that one of the authors most associated with niche-construction theory — Kevin Laland, whose work on cultural transmission appears later in this unit — is also a leading advocate for the broader theoretical reform. We can use the well-supported empirical claim (organisms alter their selective environments, sometimes durably) without committing to the contested claim (that this overturns the structure of evolutionary theory). Natural selection is not being rejected. The beaver does not escape selection by building a dam; its behavior helps determine the conditions under which selection subsequently operates. The useful correction is narrower and secure: stop picturing organisms as passive objects molded by a world they never affect.
Niche construction changes the causal diagram from a line into a loop.
Richard Dawkins used the term extended phenotype for effects of genes that reach beyond the body — a beaver dam, bird nest, or parasite-induced change in host behavior. Niche construction and the extended phenotype overlap, but they ask somewhat different questions.
The extended-phenotype perspective begins with the organism and asks how inherited variation produces effects outside its body. Niche construction begins with the feedback and asks how organism-produced environmental changes alter development and selection. A dam can be both: an extended effect of beaver traits and a constructed niche that changes the future environment.
Neither concept implies foresight. A structure can have large evolutionary effects even when no animal understands what it is building at the level at which a biologist later describes its function.
48.4 The expensive infant
The first unit introduced the human brain as an energetically expensive organ and human childhood as an unusually long period of dependence. We return to that problem because the social niche was not merely a cultural decoration added after the brain became large. Social provisioning and care were among the conditions that made a slowly developing, highly plastic human brain viable in the first place.
Every species must solve the problem of parental investment, but species solve it differently. In most mammals, mothers provide most direct care; biparental care is much more common among birds. Some offspring become independent quickly, whereas others require prolonged protection and provisioning. Human children occupy an extreme part of that continuum. They are born neurologically immature, remain unable to obtain all of their own food or manage local dangers for years, and continue learning difficult subsistence and social skills well beyond weaning. The same long development that permits extensive plasticity also creates a long period during which a child consumes resources it cannot yet replace.
The phrase cooperative breeding is used for systems in which individuals other than the mother contribute to offspring care. Applied to humans, the term should not imply one universal family structure or a colony-like reproductive division of labor. Assistance varies enormously across ecologies and histories. Fathers, grandparents, older siblings, other relatives, and unrelated group members may provide food, carrying, protection, supervision, or instruction; in some settings several of these contributions are substantial, and in others they are sparse or distributed differently. The arrangements may be affectionate, reciprocal, obligatory, unequal, or coercive. What recurs is not a single social blueprint but development within a network wider than the mother–infant pair [@hrdy2016; @burkart2025].
This evidence supports a coevolutionary hypothesis rather than a simple historical sequence. Shared food and care can reduce the immediate cost of prolonged dependency. Prolonged dependency creates more time to learn from several individuals. Socially acquired knowledge can later improve foraging, food processing, childcare, cooperation, and technology. Those gains make it easier for a group to support another cohort of slow-developing children. The social niche subsidizes learning, and learned competence helps maintain the social niche.
The direction of causation cannot be reduced to “large brains caused cooperation” or “cooperation caused large brains.” Diet, food sharing, life history, social tolerance, alloparental care, cultural learning, and brain development altered one another over long timescales. Cooperative breeding is an influential candidate for an early catalyst in that feedback, not a demonstrated single trigger [@burkart2025]. The broader claim is firmer: human development normally occurs within systems that redistribute energetic and informational costs across people and time.
This widens the energetic argument made in Chapter 2. Maternal metabolism constrains gestation and lactation, but maternal energy is embedded in a social economy. A mother can forage while another person watches a child. A child can consume food acquired or processed by someone else. Storage and cooking can move calories across time; sharing can move them across bodies. Teaching can transfer information whose independent discovery would be dangerous or slow. Social organization therefore changes the resources available to a developing nervous system.
The human brain, in this sense, is an individual organ whose ordinary construction presupposes social infrastructure. That does not make the group an organism, nor does it make care harmonious. It means that the developmental budget of one child is often balanced by the work and knowledge of several other people.
Cooperative breeding raises a classic evolutionary problem. If natural selection favors traits that increase reproductive success, why spend time and energy helping to raise another individual’s offspring?
There is no single answer. Help can benefit relatives who share genes, as formalized by Hamilton’s theory of inclusive fitness [@hamilton1964]. It can produce immediate mutual benefits when several individuals do better together than alone. It can be exchanged over time, the logic Robert Trivers developed as reciprocal altruism [@trivers1971]. Care can also improve reputation, preserve valuable partners, reduce the future burden on helpers, or be enforced through social expectations.
Trivers belongs in this unit because he helped make social behavior an evolutionary problem rather than a moral exception to evolution. His work on reciprocal altruism, parental investment, and parent–offspring conflict showed that care and cooperation can evolve without assuming that natural selection has made individuals selfless [@trivers1971; @trivers1974]. Cooperation is often real; so are conflicts over its costs. The same body of theory predicts that a mother and her infant, or two parents, or a helper and the young it tends, will frequently disagree about how much should be given and by whom.
We return to this logic when we consider social understanding. For now the important point is that alloparental care need not rest on one mechanism. Human social systems combine kinship, reciprocity, mutual dependence, reputation, coercion, affection, and culturally prescribed obligation.
48.6 Learning escapes the individual
Genes carry information across generations, but they are slow. Natural selection can alter a population over many generations, yet an individual animal must survive the particular conditions it encounters during its own life. Learning provides a faster route. A young rat can discover which foods are safe; a bird can learn where fruit ripens; a predator can improve its technique without waiting for a genetic change in the species.
Individual learning, however, has an obvious weakness. Discovery is expensive. An animal that learns by trial and error must pay for its errors: the wrong food may be poisonous, the wrong route may end at a cliff, the wrong prey-handling technique may produce a sting, bite, or broken tooth. Even successful exploration consumes time and energy. And when the animal dies, much of what it learned disappears with it.
Social learning allows one animal to use information acquired by another. An individual can observe where companions forage, which foods they avoid, how they open a shell, where they migrate, or which call signals danger. The learner may watch the behavior directly, follow the path it leaves, or respond to a changed environment the demonstrator produced. In each case, information moves between individuals without passing through DNA. This is not a rare trick confined to apes. Social learning occurs in insects, fish, birds, and mammals, and the evidence has become broad enough that the existence of animal culture is no longer a serious point of dispute, though the complexity and mechanisms differ greatly among species [@whiten2021].
Consider migration. It is tempting to describe a migration route as an inherited program written into the nervous system, and in many species genetic information clearly contributes to when and in what general direction an animal moves. Yet the route can also be a social inheritance. In a reintroduced population of whooping cranes, younger birds improved their migratory performance when they traveled with older individuals; experience residing in the group helped preserve and refine a route across years [@mueller2013]. The landscape did not change, and the cranes’ genes did not change on the timescale of the study. What changed was the information available in the flock.
The flock became part of the navigation system.
A similar point applies to chimpanzee communities, which differ in socially maintained behaviors — methods of extracting termites, cracking nuts, grooming, displaying, using leaves — that cannot be explained simply by local ecology or genetic difference [@whiten1999]. No one behavior is equivalent to human technology, but that is not the relevant comparison. The important fact is that a young chimpanzee develops within a community that already has a repertoire. The local group changes what the youngster is likely to notice, practice, and eventually master. Social learning therefore modifies the developmental niche: the novice does not confront all possible actions equally. Other animals direct attention toward some objects, make some techniques available for observation, tolerate some mistakes, and respond differently to successful and unsuccessful behavior. The group reduces the space through which the learner must search.
That can be a great advantage. It can also propagate errors. Copying others is useful only when others possess information worth copying, and a group can preserve an outdated route, an inefficient technique, or an arbitrary preference long after the original reason has disappeared. Social learning trades some of the cost of individual discovery for a new risk: dependence on the history and judgment of the group.
This is the beginning of culture.
The word culture often evokes literature, religion, music, cuisine, law, and the arts. Biologists use it more broadly. A behavior is usually described as cultural when it is acquired at least partly through social learning and becomes shared or persistent within a group.
That definition excludes several look-alikes. A behavior is not necessarily cultural merely because many animals perform it; all members of a species may build similar nests because of shared genes and similar environments. Nor does every difference between groups demonstrate culture; two populations may behave differently because their habitats differ.
The strongest evidence combines several observations: individuals acquire the behavior from others, the behavior spreads through social networks, local traditions persist, and ecological or genetic explanations are insufficient. Under this definition, culture does not require language, symbolic meaning, or conscious teaching. It requires socially transmitted information.
Using the same word for whale song and constitutional law does not imply that they are equivalent. It identifies a real continuity — information moving socially across individuals and generations — while leaving enormous room for differences in scale, representation, and organization.
48.7 Teaching changes the learner’s world
Social learning does not require teaching. A young animal may simply watch what an experienced animal happens to do. Teaching is more demanding, because the experienced individual changes its behavior in a way that helps the novice learn, usually at some cost or without an immediate benefit to itself.
Wild meerkats provide a concrete example. Their pups must learn to handle scorpions — nutritious prey capable of delivering a painful and dangerous sting. Adults do not merely drop identical prey in front of pups of all ages. They adjust what they provide: younger pups are more likely to receive dead or disabled scorpions, older pups increasingly intact ones. The adult’s behavior scaffolds the learner’s exposure to danger, and experiments that altered the begging calls heard by adults shifted the prey they supplied [@thornton2006].
The lesson is larger than meerkats. A teacher can alter the difficulty of the environment so that a learner encounters a sequence of manageable problems. In control-theory language, the novice is not simply adapting to a fixed task; another organism is changing the task as the novice changes. Human caregiving is saturated with this kind of scaffolding. Caregivers hold an object where an infant can reach it, exaggerate a gesture, repeat a word, break a procedure into steps, prevent a dangerous error, and gradually withdraw assistance — much of it without any formal lesson. The social niche is continuously adjusted around the developing child.
Teaching can also transmit what observation alone cannot reveal. An action may have a hidden causal structure: a plant may be safe only after soaking, a tool may need to be held at an angle that is difficult to infer from its outcome, a ritual may matter because of a rule rather than any visible physical effect. Demonstration, correction, and eventually language make these otherwise opaque relationships learnable. Once survival depends on knowledge too difficult, dangerous, or time-consuming for each individual to rediscover, selection can favor both greater dependence on social learning and social arrangements that make learning reliable. The niche becomes richer in information while the individual becomes more dependent on inheriting it.
That dependency is not a defect. It is the price of entering a more capable system.
48.8 Culture did not begin with humans
For much of the twentieth century, culture was treated as a bright line separating humans from other animals. The line has not survived observation. Songbirds maintain local dialects. Humpback whale songs change and spread across ocean basins. Killer-whale groups possess distinctive vocal and foraging traditions. Chimpanzee communities differ in tool use and social conventions. Fish copy routes and food choices. Bumblebees learn flower-handling techniques from demonstrators. Culture, in the broad biological sense, has appeared repeatedly because social information is useful wherever the world is variable and costly to explore.
This continuity matters for an evolution-first book. Human culture did not arrive from nowhere. Its prerequisites were assembled gradually: attention to other individuals, tolerance of proximity, memory for observed actions, motivation to copy, sensitivity to success, prolonged development, and opportunities for repeated interaction. Different species possess different subsets and elaborations of these capacities. Saying that animals have culture does not mean that a bee colony has jurisprudence or that whale song is a language in the human sense. It means that the social transmission of behavior is evolutionarily old and taxonomically widespread. The question is no longer whether humans alone possess culture. It is what changed the scale, fidelity, flexibility, and reach of cultural inheritance in our lineage.
One long-standing answer was cumulative culture.
48.9 The ratchet
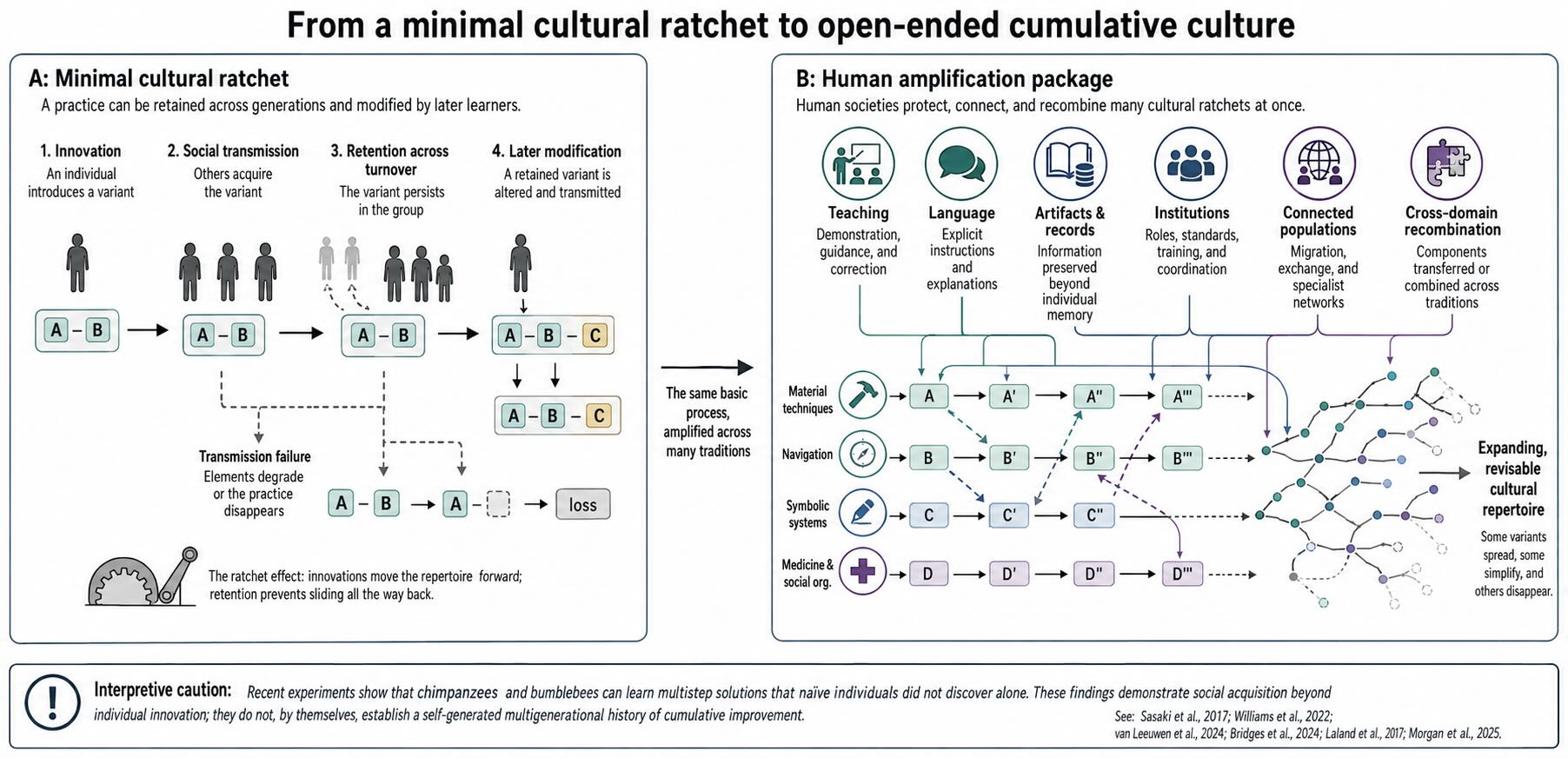
Suppose one animal discovers a better way to open a nut. Others acquire the technique. A later individual modifies it, and that modification is transmitted in turn. If later learners preserve enough of what came before, the practice can become more effective or more complex than anything a novice would be likely to invent alone. Cultural change has accumulated.
Michael Tomasello and others described this as a ratchet effect. Innovations move a repertoire forward, while social transmission prevents it from sliding all the way back. The metaphor captures something undeniable about human life. No person begins by rediscovering controlled fire, metallurgy, algebra, anesthesia, or semiconductor fabrication. Each generation enters a world already containing solutions, conventions, tools, and institutions produced by people it will never meet.
For a time, cumulative culture was treated as an all-or-none human distinction. Comparative evidence now makes that boundary harder to defend. Pairs of homing pigeons can pass routes through experimental chains, with later pairs sometimes finding more efficient paths [@sasaki2017]. Socially learned song elements in a wild population of Savannah sparrows changed over successive generations in ways that increased their behavioral efficacy [@williams2022]. Recent experiments also show that chimpanzees and bumblebees can acquire multi-step solutions from demonstrators after naive individuals failed to discover the full solution during individual exposure [@vanleeuwen2024; @bridges2024].
Those results matter, but they should not be asked to prove more than they do. The chimpanzee and bee experiments demonstrate social acquisition of behavior beyond what the tested individuals independently innovated under the experimental conditions. They do not demonstrate a long historical sequence of improvements generated and preserved by the animals themselves. Pigeon routes and bird songs accumulate within restricted domains; they are not thereby equivalent to an expanding technological system. The useful conclusion is not that human and nonhuman culture are the same. It is that accumulation has components and degrees, and that no single small-scale success provides a clean species boundary.
What makes human culture so unusually powerful is probably a package rather than one magic mechanism. Transmission fidelity matters because a fragile, multi-step procedure decays if every learner reconstructs it too loosely. Imitation, demonstration, teaching, language, and norm enforcement can all stabilize transmission [@laland2017book]. Recombination matters because a solution developed in one domain can be imported into another. External storage matters because artifacts, diagrams, writing, and digital records preserve information beyond biological memory. Population structure matters because specialists, teachers, migrants, and connected groups determine which variants meet and which skills survive. Institutions matter because they assign roles, enforce standards, and coordinate people who never interact face to face.
The result is not merely a better ratchet. Human groups build systems that protect, compare, recombine, and deliberately modify many ratchets at once. A knot used in fishing can enter surgery. A counting practice developed for exchange can become mathematics. A tool creates materials for later tools; a school preserves procedures that no teacher invented; a laboratory allows hundreds of partial experts to extend a result no one person fully understands. The space of possible cultural combinations grows as culture grows [@morgan2025].
Open-ended cumulative culture is therefore a useful description of the human pattern, provided it is treated as an empirical profile rather than a metaphysical boundary. Humans are exceptional in the scale, range, institutional support, and cross-domain recombination of cultural inheritance. Whether one ingredient deserves priority remains unsettled. What is clear is that the inheritance system resides in relationships among learners, teachers, artifacts, and networks—not in copying fidelity alone.
In the bumblebee experiment, the reward was hidden behind a two-step mechanism. A bee first had to move one part of the apparatus away from the target and then perform a second action to gain access to sugar. Naive bees were given extensive opportunities but did not solve the full sequence independently. Bees that watched a trained demonstrator could acquire it [@bridges2024].
This result is interesting because the observer learned a behavior beyond what members of the tested population spontaneously invented under the experimental conditions. It weakens the claim that only humans can socially acquire solutions they cannot individually generate.
It does not show that bumblebees possess human-like cumulative culture. The experimenters trained the initial demonstrators; the bees did not produce a long historical sequence of improvements, teach symbolically, or recombine the technique with an expanding technological repertoire. A good comparative result should sharpen a distinction rather than force a choice between “just like us” and “nothing like us.”
48.10 The network remembers
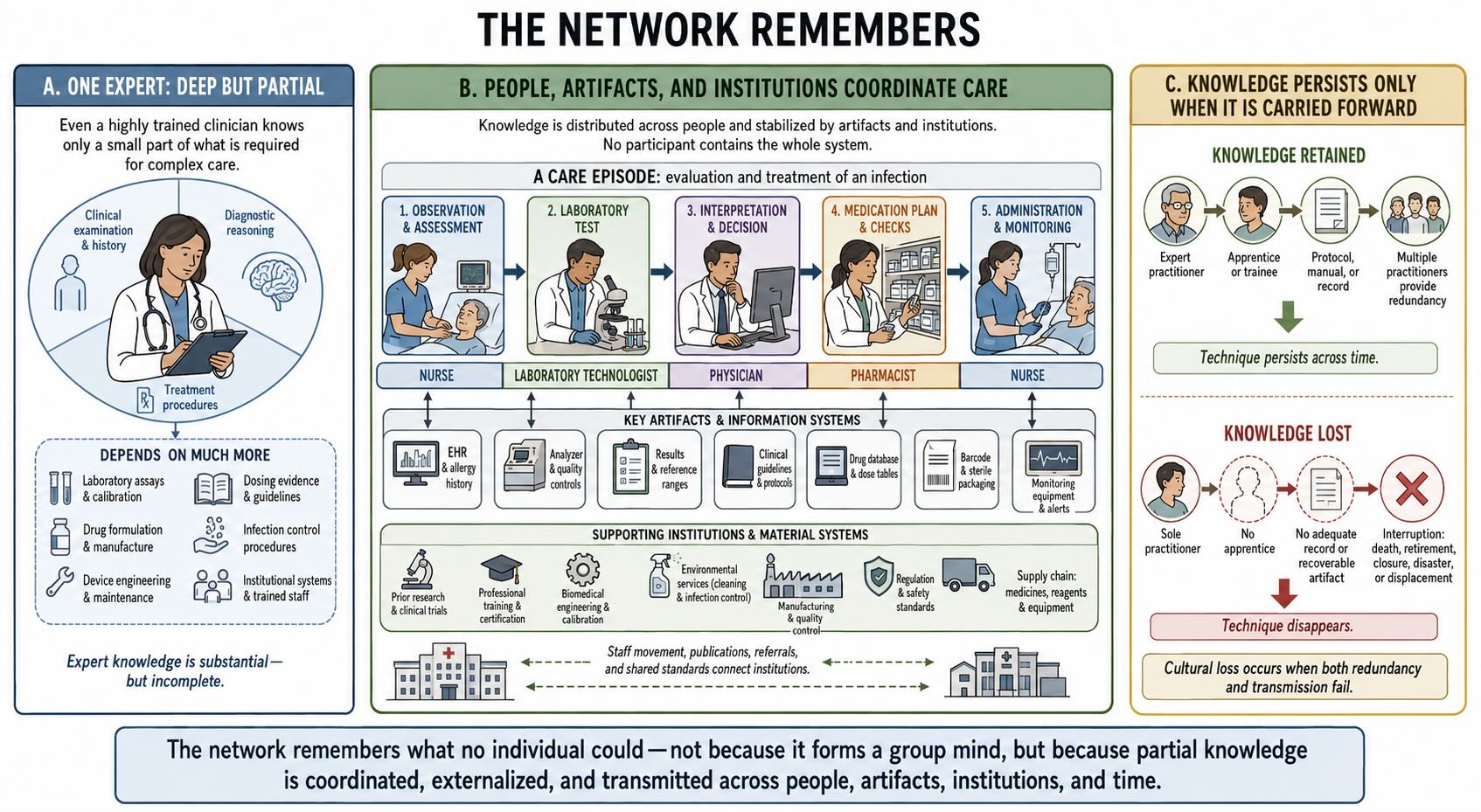
Culture is often discussed as though it were a property stored inside each person’s head. Some of it is. You know words, routes, customs, facts, and techniques that you acquired from others. But no individual contains more than a small fraction of the culture on which that individual’s life depends.
Consider an ordinary hospital. A patient survives because of knowledge distributed among physicians, nurses, pharmacists, laboratory technologists, engineers, cleaners, manufacturers, statisticians, and generations of investigators who never entered that building. The relevant information also resides in instruments, dosage tables, sterile packaging, electrical standards, supply chains, protocols, laws, and computer systems. Remove enough of that network and the expertise of any one person becomes insufficient.
The same is true of less technological societies. Knowledge of landscapes, seasons, plants, animals, social obligations, food processing, and childcare may be distributed by age, sex, experience, family, or specialty. Different people know different things. Mobility and marriage connect local groups, allowing useful practices and materials to move beyond the band in which they originated. Models and empirical work suggest that the multilevel social networks characteristic of hunter-gatherers can accelerate the recombination and accumulation of cultural information [@migliano2020].
This is one reason population structure matters. A large population does not automatically produce more culture, and a small population does not automatically lose it. What matters includes who interacts, how often people move among groups, whether novices can observe experts, how knowledge is divided, and whether rare skills have enough practitioners to survive interruption. A culture can lose a technique without anyone deciding to abandon it, simply because the last people who know it die before transmitting it.
The cognitive unit is therefore sometimes larger than the individual without becoming a mystical group mind. A navigation route may be maintained by a flock. A tool tradition may be maintained by a community. A scientific field may preserve methods that no member understands in their entirety. Information is distributed across brains and stabilized by relationships and artifacts.
The network remembers what no individual could.
This returns us to allostasis. A network can pool not only calories but knowledge. The expert who recognizes a medicinal plant, predicts a storm, repairs a pump, or diagnoses an infection changes the regulatory options available to everyone connected to that expert. Cultural specialization allows individuals to depend on solutions they do not personally understand. The gain is enormous capacity; the cost is systemic dependence. A person living in a modern city may know almost nothing about obtaining clean water, generating electricity, growing grain, or manufacturing antibiotics, yet those resources arrive because the cultural niche contains institutions that organize the necessary knowledge and labor. Human adaptability often belongs less to a versatile isolated brain than to a brain able to enter, trust, navigate, and contribute to a versatile network.
This is the murder of brains in its fully human form: a description of where the relevant cognition actually lives. The single brain is necessary but radically insufficient. What keeps a human alive — what carries the calories, the care, and the accumulated knowledge across the long dependency of childhood and the longer interdependence of adult life — is a population of brains coupled by relationships, language, and artifacts, inheriting a constructed niche from the brains that came before.
48.11 Culture as an inheritance system
We can now identify several ways in which the past reaches into the present. Genes are inherited through reproduction. Constructed environments can persist as ecological inheritance. Socially learned information can persist as cultural inheritance. These channels overlap, but they are not interchangeable.
Cultural information can travel vertically, from parents to offspring; horizontally, among members of the same generation; or obliquely, from unrelated older individuals to younger ones. It can cross group boundaries, spread epidemically through a population, or disappear within a single generation. It may be carried by behavior, speech, song, gesture, artifacts, images, institutions, or written records. This makes cultural inheritance faster and more flexible than genetic inheritance. A useful technique can spread among thousands of adults who share no recent ancestor. A person can acquire several incompatible traditions, reject part of what was taught, or deliberately import a practice from another society. Cultural lineages branch, merge, borrow, and revive.
The word inheritance should therefore not imply exact copying. DNA replication is itself imperfect, but cultural transmission is far more reconstructive. Learners attend selectively. They infer goals. They simplify, embellish, misunderstand, and combine. A story retold is not a molecular duplicate of the story heard; an apprentice may reproduce the outcome while changing the method; a law can remain on the books while its interpretation changes. This is exactly why fidelity of imitation matters so much, and why the cases where it is high — opaque procedures faithfully copied — are the ones that let culture accumulate rather than drift.
Nevertheless, persistence is real. Languages, recipes, rituals, technologies, and institutions can outlive the people who introduced them, and their effects on development and behavior can be as concrete as those of a physical structure. Artifacts make this especially visible. A stone tool embodies choices about material, shape, and sequence. A road channels later movement. A house organizes privacy, temperature, family interaction, and exposure to pathogens. A written text allows one person’s marks to alter another person’s nervous system centuries later. The object is not alive, but it carries forward constraints and possibilities created by prior minds.
Culture is thus both information and environment. It tells people what to do, and it builds worlds in which some actions become easy, expected, compulsory, or nearly impossible.
48.12 Culture is not the opposite of biology
At this point the chapter reaches a distinction that has done both useful and damaging work. Human behavior is often divided into two explanatory domains. Biology is assigned genes, bodies, selection, and what humans share with other animals. Culture is assigned language, institutions, history, meaning, morality, art, and choice. The division protects important forms of explanation. A poem is not explained by sequencing the poet, and a legal system cannot be derived from inclusive fitness. Historical contingency, interpretation, power, and deliberate action are genuine causes in human affairs.
The mistake begins when explanatory difference is turned into ontological separation.
Culture is produced, learned, remembered, and enforced by living organisms. It depends on perception, movement, motivation, memory, development, and social interaction. Its forms are transmitted through speech, gesture, artifacts, practices, and institutions. In that broad sense, culture belongs within natural history. Whale songs and chimpanzee tool traditions do not cease to be biological phenomena because they are socially learned, and human traditions do not require a nonbiological substrate merely because they become symbolic, normative, or extraordinarily complex.
That claim does not mean that a biological description is sufficient for every question. Knowing that reading recruits visual and language networks does not tell us what a novel means. Knowing that an institution is maintained by reward, punishment, and social learning does not tell us whether the institution is just. Levels of explanation can be physically compatible without being interchangeable. The architecture-first position of this book rejects both a culture floating free of bodies and a reductionism that treats meaning or history as decorative language for genes.
A second ambiguity matters even more. Biological is often used as though it meant genetically specified. It does not. A scar is biological but not inherited in DNA. The cortical changes produced by literacy are biological but are not genetic adaptations for Roman letters. A child develops a brain in response to nutrition, stress, language, schooling, tools, expectations, and relationships. Those effects are embodied and measurable even when no relevant difference in allele frequency exists.
This point carries an ethical as well as scientific consequence. Finding a difference in brains does not establish that the difference is innate, fixed, or genetic. Brains are the organs through which experience has effects. If two populations encounter different languages, educational systems, hazards, diets, or social expectations, their average behavior and neural organization may differ because development has followed different histories. Putting culture inside biology therefore does not collapse culture into genes. It makes the distinction between genetic inheritance and developmental construction more important.
Human culture is also genuinely distinctive without being exempt from evolutionary continuity. Other animals socially transmit songs, routes, tool use, and conventions. Humans add an extraordinary scale of symbolic reference, explicit teaching, normative enforcement, external records, institutions, and cross-domain recombination. Some of those differences may amount to reorganizations that deserve new explanatory concepts, just as multicellularity deserves concepts not needed for a single cell. Emergent organization is not supernatural, and continuity of origin does not imply sameness of outcome.
The defensible position is therefore neither “culture is merely biology” nor “culture begins where biology ends.” Culture is an inheritance system and a constructed environment implemented by evolved, developing organisms. Particular cultural forms still require historical, social, and interpretive explanations at their own level. The process is continuous with the rest of life; the explanatory vocabulary becomes richer because the organization becomes richer.
That position lets us ask the next question with precision. How can a cultural environment alter a brain? There are at least two routes, and confusing them is one of the fastest ways to mistake a developmental effect for a genetic one.
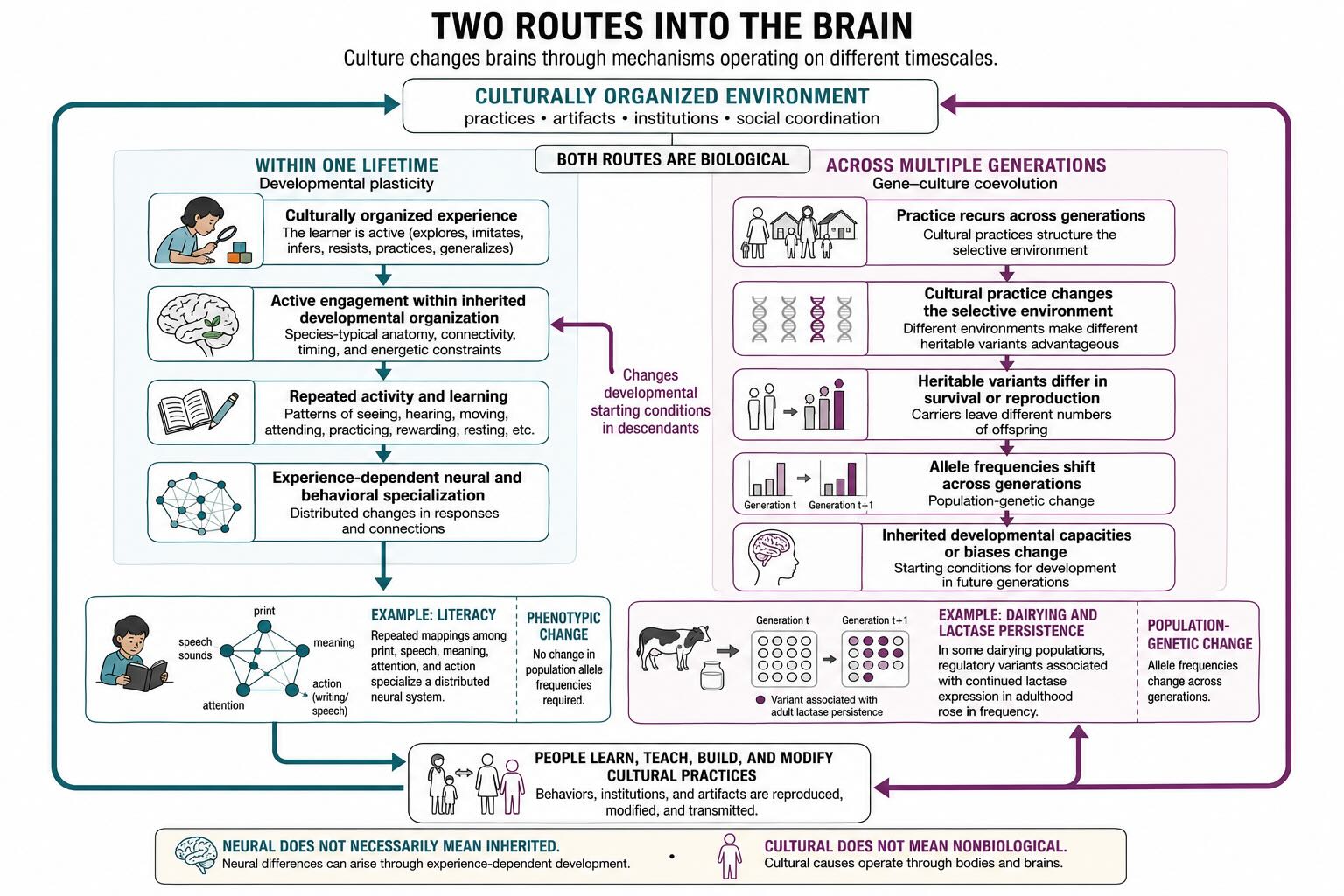
48.13 Two routes into the brain
Culture changes brains through mechanisms operating on different timescales.
Across generations, a recurrent cultural environment can change which genetic variants are advantageous. Dairying alters the value of adult lactase production; dense settlements alter pathogen exposure; new forms of food acquisition or social coordination can alter the returns on learning, anatomy, and life history. This is gene–culture coevolution. Culture supplies part of the selective environment, and genetic evolution changes the capacities and biases through which later culture is acquired.
Within a lifetime, culturally organized experience changes the phenotype produced by a developing organism. This is developmental plasticity. A child is not born genetically specified to speak English, Mandarin, Arabic, or a signed language. The nervous system develops capacities for language learning, but the local community supplies the particular contrasts, words, constructions, and conversational practices. A child is not born with a cortical region selected for alphabetic reading. Literacy repeatedly engages visual, auditory, language, attention, and motor systems until their responses and connections become specialized for a cultural invention.
Musical training, calculation, tool use, navigation, and professional expertise follow the same general route. The learner is not passive: children explore, imitate, infer, resist, practice, and generalize. Nor is the brain infinitely malleable: development proceeds within species-typical anatomy, connectivity, timing, and energetic constraints. The phenotype is constructed through an interaction between those inherited constraints and a socially organized history of activity.
Both routes are biological, and they can interact. A genetic bias can alter what is easy to learn; a learned practice can alter selection; the resulting population can then build a different developmental environment. But the routes are not interchangeable. Gene–culture coevolution changes the distribution of inherited variants across generations. Developmental plasticity changes one organism during its lifetime without requiring any change in gene frequency.
This distinction prevents two opposite errors. The first is geneticizing a neural difference: assuming that because a group difference can be measured in the brain, it must have been inherited genetically. The second is disembodying cultural causation: assuming that calling a difference cultural removes physiology and development from the explanation. A school changes brains through repeated patterns of seeing, hearing, moving, attending, practicing, sleeping, eating, reward, and stress. Cultural causation reaches the nervous system through bodies.
The next chapter will make the developmental route concrete. Language is an inherited feature of the social environment that specializes perception and reorganizes a distributed neural system during childhood. The genetic route is clearest when a cultural practice changes natural selection itself.
48.14 Gene–culture coevolution
The most direct challenge to the biology-versus-culture divide comes from the first route — cases in which cultural behavior changes genetic selection.
Genes influence the capacities and biases through which culture is acquired. They help construct sensory systems, learning mechanisms, social motivations, vocal anatomy, and the prolonged development on which cultural dependence rests. Culture then changes diet, mating, disease exposure, movement, workload, fertility, and survival. Those changes alter which genetic variants are advantageous. The two inheritance systems begin to coevolve [@richerson2010].
The classic example is adult digestion of lactose. All mammalian infants produce lactase, the enzyme that breaks the milk sugar lactose into absorbable components. In most mammals, and in many humans, lactase production declines after weaning — which makes biological sense in a world where milk is available only during infancy. Once some human populations began herding and milking animals, however, the nutritional environment changed. Milk became available to older children and adults, and genetic variants that maintained lactase production into adulthood — lactase persistence — could now provide an advantage.
The sequence matters. Dairying was a cultural innovation; it created a new food source and thereby changed the selective value of existing genetic variation. Ancient-DNA evidence indicates that early European farmers used dairy products long before the now-common European lactase-persistence allele became frequent [@burger2007]. Culture changed first; genes followed. Even this famous example is less tidy than textbook versions suggest. Milk consumption alone does not fully explain the timing and geographic spread of lactase persistence; recent work argues that episodes of famine and pathogen exposure may have made the consequences of drinking fresh milk without digesting lactose especially dangerous, strengthening selection under particular historical conditions [@evershed2022]. The important point survives the complication: a culturally created dietary niche altered human genetic evolution, but the strength of that alteration depended on ecology, demography, and disease.
This is the loop in its most rigorous form. A learned practice changed the environment; the changed environment changed selection on genes; and the resulting genetic change was itself only legible in the context of a particular constructed niche. Gene–culture coevolution is not a metaphor or an analogy to biological evolution. It is biological evolution, with culture supplying part of the selective environment.
The answer depends on what one means by Darwinian.
If the term requires blind variation, particulate inheritance, and faithful vertical transmission from parent to offspring, then much cultural change is not Darwinian in the strict genetic sense. Cultural variants can be deliberately designed. They pass horizontally and obliquely. Several sources can blend. Learners reconstruct rather than copy. Acquired changes are routinely transmitted.
If the term refers more generally to descent with modification and differential persistence among inherited variants, then cultural evolution is plainly Darwinian enough to support useful theory. Technologies form lineages. Languages diverge and borrow. Practices spread through biased social transmission. Some variants proliferate because they work; others because powerful institutions enforce them; still others by chance.
The safest conclusion is neither that culture works exactly like genes nor that it is exempt from evolutionary analysis. Cultural inheritance is an evolutionary system with unusual transmission rules [@creanza2017; @richerson2010]. Its differences from genetics are part of the subject, not objections to the subject.
In everyday speech, evolve often means improve. Biological evolution has no such guarantee, and cultural evolution does not either.
A cultural trait can spread because it benefits its users, because it benefits elites who enforce it, because it exploits attention, because it is easy to remember, or because it happens to be attached to something else that is useful. A practice can persist after its original function is gone. A system can become more complex while making many lives worse.
The evolutionary claim is that culture changes through inherited variation and differential persistence. It is not that history moves upward toward wisdom.
48.15 The brain the niche paid for
Chapter 2 treated human encephalization in detail: the allometric problem, the fossil trajectory, the metabolic cost of neural tissue, and the competing explanations for why that cost was favored. The final unit does not need to repeat that evidence. It needs to place the evidence inside the reciprocal loop developed here.
The human brain is expensive not only to operate in adulthood but to construct over a prolonged childhood. Its expansion therefore required more than a benefit. It required an ecology and life history capable of supplying energy reliably enough, and for long enough, to pay the developmental bill. The influential expensive-tissue hypothesis proposed that improved diet allowed energy to be reallocated from a relatively costly gut toward a larger brain [@aiello1995]. Comparative tests have not supported a simple universal tradeoff between gut and brain size, so the hypothesis is best retained as one part of a larger energetic argument rather than as a settled accounting identity [@navarrete2011].
The larger argument involves a linked set of changes. Higher-quality and mechanically or thermally processed food can increase net energy return. Food sharing and alloparental care can move that energy toward mothers and dependent children. A longer juvenile period can increase the amount learned before independent reproduction. Social learning can make the return on that prolonged development larger because the learner receives knowledge that no individual could cheaply rediscover. Improved tools and accumulated expertise can then increase the reliability of food acquisition and care. Each component changes the value and affordability of the others.
The fossil record establishes broad changes in endocranial volume and shape across hominin evolution, but it cannot assign those changes to one social or cultural cause [@holloway2015]. Nor can comparative correlations among brain size, diet, group structure, and life history provide a clean historical sequence. The field remains divided about the relative contributions of ecological challenge, foraging, cooperative care, social competition, cultural dependence, and developmental constraints. That uncertainty is not a reason to retreat to a brain-first story. It is a reason to prefer a coevolutionary one.
The most defensible claim is about the form of the explanation. Hominin brain expansion was not a completed biological event followed by the late appearance of culture. Diet, life history, care, learning, technology, and neural development changed in feedback. A richer niche made a costly brain more affordable. A slowly developing brain increased the capacity to acquire and extend that niche. The returns were realized not by a solitary clever individual but by individuals embedded in systems of provisioning and inherited knowledge.
This is the same causal shape shown by the hive and the mound, but not the same organization. Bees and termites achieve colony-level regulation through evolved divisions of labor and local interactions. Humans add flexible learning, symbolic communication, institutions, and cumulative artifacts. The analogy is the loop, not a claim that a human society is one organism.
The phrase the brain the niche paid for should therefore be read carefully. A niche does not write a check, and evolution does not plan an investment. The phrase names a constraint: an organ with a large energetic and developmental cost can persist only where the ecological and social returns offset that cost. In the human case, many of those returns existed only because other people and inherited practices were already part of the system.
Brains do not fossilize. Paleoneurologists therefore work mainly with endocasts—natural or reconstructed impressions of the cranial cavity. Endocasts can estimate endocranial volume and sometimes preserve broad features of external shape, vascular impressions, or major sulcal landmarks [@holloway2015].
They cannot reveal neurons, microcircuits, neurotransmitters, white-matter connectivity, or the computations performed by living tissue. Even surface landmarks may be faint, distorted, or disputed. A change in cranial capacity is therefore much firmer evidence than a claim that one fossil possessed a modern language network, a particular cognitive faculty, or a named cortical area with a modern function.
The limitation does not make paleoneurology uninformative. It defines the level at which its evidence is strongest. Endocasts constrain the size and gross organization that any evolutionary account must explain. Comparative anatomy, archaeology, genetics, life history, and behavior must supply the rest. As throughout this book, the responsible inference is the one that preserves what the method can show without turning anatomy into an unobserved psychology.
48.16 Cooking and the metabolic budget
One cultural practice deserves separate attention because it shows with unusual clarity how learned behavior can enter the body’s energy economy.
Food processing occurs outside the gut. Pounding, grinding, slicing, fermenting, and heating disrupt tissues and alter the work required to chew, digest, and absorb them. Experimental work shows that mechanical and thermal processing can increase the energy organisms obtain from a given food and reduce some of the costs of handling it [@carmody2011; @kraft2021]. Cooking can also detoxify some foods, improve storage, and make resources shareable in forms that children and older adults can use.
The historical argument is more difficult. A strong version proposes that habitual cooking relaxed constraints on teeth, jaws, gut, and feeding time and thereby helped make major hominin brain expansion possible. The proposal is plausible and influential, but the chronology of controlled fire and routine cooking remains too uncertain to assign cooking a single decisive place in the sequence. Meat eating, tool-assisted processing, food sharing, climate, and changes in life history also altered energetic return. Archaeology rarely tells us how regularly a fire was used or what proportion of a population’s diet was cooked.
The chapter does not require the strongest chronology. The secure point is already consequential: once culturally learned processing changed the net return, safety, timing, or distribution of food, it changed the developmental environment of bodies and brains. Knowledge about how to transform food could be transmitted socially, stored in tools and routines, and shared across people who contributed different parts of the labor.
The loop is therefore direct:
learned processing → altered food → altered energy return and availability → altered developmental and selective possibilities
Culture entered the metabolic budget. That statement does not tell us when the first regular hearth appeared or how much of encephalization it explains. It identifies a mechanism by which a socially inherited practice can become part of the biological economy that supports a brain.
48.17 Constructed niches create new problems
Niche construction can sound like an evolutionary success story: animals improve their surroundings, and humans improve them more. That is not what the concept means.
Organisms alter environments, but they do not control all the consequences. A niche can buffer one disturbance while amplifying another. It can benefit one individual and burden another. It can produce dependence on conditions that later disappear. It can make short-term regulation easier while creating long-term instability.
Agriculture is the obvious human example. Cultivation and animal domestication increased the amount of food that could be produced in a given area and supported larger, denser populations. Stored surpluses buffered seasonal variation, and settled communities enabled specialization and institutions that would have been difficult to maintain in mobile groups. The same niche introduced new vulnerabilities. Dependence on a small number of crops made harvest failure catastrophic. Density and proximity to domesticated animals altered infectious-disease ecology. Stored food attracted pests and made unequal control of surplus possible. Sedentary labor changed bodies and workloads. Institutions that coordinated production could also impose taxation, hierarchy, forced labor, and war.
The allostatic language helps here. A regulatory solution is always organized around particular variables and timescales. A granary can stabilize calories across winter while increasing the political power of whoever controls the granary. Irrigation can buffer rainfall while salinizing soil. A city can deliver clean water to millions while making them dependent on pumps, electricity, chemicals, and institutions they do not personally control.
Modern medicine offers the same mixed lesson. Antibiotics transformed bacterial infection from a common cause of death into a treatable condition. Their use also created a powerful selective environment favoring resistant organisms — the cultural niche changing microbial evolution with extraordinary speed. We solved one regulatory problem and created another.
This is not an argument against construction. There is no return to an unconstructed human life. It is an argument against confusing control with mastery. Every intervention enters a system with feedback, delay, conflict, and unanticipated effects. Constructed niches also produce path dependence: once roads, cities, legal systems, or technologies are in place, later choices are made within the constraints they establish. A technically inferior standard can persist because too many other systems depend on it. A harmful institution can reproduce itself by shaping the incentives and beliefs of those raised within it. The inherited niche narrows the space of imaginable alternatives.
This also returns us to a point made earlier about intention. Humans can deliberately alter a tool, a law, or an institution because they imagine a better outcome, and that capacity for foresight is real. But it is local and limited. Agriculture was not invented in order to create lactase-persistence alleles, epidemic disease, states, or modern property law. Antibiotics were not developed to evolve resistant bacteria. Intentional actions enter feedback loops whose long-term consequences escape the intentions that began them. Culture gives evolution foresight only in a narrow and local sense; it does not teleport the result outside nature.
Animals create niches that modify animals. Humans create cultural niches that modify humans — often in ways no human planned. Sometimes the modification is liberating. Sometimes it is constraining. Usually it is both.
Living organisms are far-from-equilibrium systems. They maintain local organization by taking in usable energy and matter and releasing heat and waste. This is fully compatible with the second law of thermodynamics: local order is sustained through energy flow while entropy increases in the larger environment.
Some physicists have asked whether driven matter can acquire forms of organization that resemble adaptation. Jeremy England’s work on dissipative adaptation, for example, explores conditions under which driven systems become organized into states that absorb and dissipate work reliably [@england2015]. The proposal is provocative because it asks whether life-like organization rests on regularities more general than biology.
The levels should not be collapsed. A flame, a hurricane, and an organism all dissipate energy, but only the organism belongs to a lineage with heredity, variation, and differential reproduction. Thermodynamics constrains what living systems can do and may help explain how organized states can arise. It does not replace evolutionary, developmental, neural, or cultural explanations. Beaver dams, teaching, reciprocal altruism, and written language require causal histories and architectures that are invisible at the level of energy flow alone.
My inclination is to say that matter in open systems can acquire history, and that heredity makes that history progressively more consequential. Genes preserve one kind of history; nervous systems add learning; culture adds socially distributed inheritance. That is a philosophical synthesis, not a settled physical law. The secure claim is narrower: every level in this book remains physical, but lower-level compatibility is not the same as explanatory sufficiency.
48.18 Coda: worlds that build brains
We began the book by asking why an animal should have a brain. The first answer was movement: a nervous system links sensation to action so that a body can remain alive in a variable world. The answer then widened. Brains regulate internal resources, choose among competing actions, learn from consequences, and prepare for disturbances before they become catastrophic. They are embodied control systems with an evolutionary history and an energetic cost.
This final unit adds the outer loop. Bodies do not merely move through environments; they alter them. Some alterations persist and become part of the developmental or selective conditions encountered by later organisms. Social animals also construct relationships, divisions of labor, systems of care, traditions, and institutions. Those arrangements redistribute calories, danger, work, and information. In humans, cultural inheritance allows the constructed niche to accumulate across generations and to contain more knowledge than any individual could discover or understand.
The causal sequence is not a ladder away from biology. Organisms act in environments. Their actions change those environments. Persistent changes affect later development and selection. Social learning lets acquired information survive the individual who first obtained it. Artifacts and institutions stabilize some of that information; language moves it; connected populations recombine it. The inherited niche then feeds back onto bodies, brains, genes, and future culture.
The human brain is therefore neither the isolated cause of culture nor a passive surface on which culture writes. Brains make culture through embodied action, and culture helps make brains by organizing energy, experience, learning, and selection. The arrows run both ways. That is the book’s architecture-first answer to a temptation that will recur in the next two chapters: do not begin with a special human faculty and search for its box. Begin with the adaptive problem, the older systems available to solve it, and the developmental and social niche in which those systems are assembled.
Chapter 49 applies that strategy to language. Speech and sign will not be treated as outputs of a detached language organ. They are ways of acting on other agents—coordinating attention, transferring knowledge, repairing misunderstanding, and preserving information beyond one brain. Their neural organization reflects the integration of older auditory, visual, motor, memory, and social systems with a cultural inheritance that every child must reconstruct.
Chapter 50 takes the final step. When survival depends on provisioning, teaching, exchange, reputation, and cooperation, other agents become among the most consequential features of the environment. Their behavior cannot always be predicted from visible movement alone. Their goals, knowledge, and mistaken beliefs may have to be estimated, and they may be estimating yours in return. The control system has encountered an object that is itself a control system.
The opening honey bee now carries the argument one last time. A bee cannot be understood completely outside the hive that bees built. A human brain cannot be understood completely outside the social and cultural world made by earlier brains. The analogy has limits: human groups contain overlapping memberships, explicit conflict, individual reproduction, and institutions that can be resisted or redesigned. But the reciprocal principle survives the differences.
Animals create niches that modify animals. For humans, the most consequential part of that niche is other people.
This overview has crossed several levels of explanation, so the final distinctions matter.
We are confident that: organisms alter environments in ways that change development and selection; persistent organism-produced changes can function as ecological inheritance; social learning and animal culture are widespread; human children undergo unusually prolonged provisioning, protection, and learning; culturally transmitted practices can alter genetic selection; food processing changes energetic return; and every human brain develops within a social and cultural environment built before that individual was born.
We have good reason to think that: cooperative care, food sharing, prolonged childhood, social learning, and cultural accumulation participated in feedback with hominin brain expansion; human cumulative culture depends on a package that includes reliable transmission, teaching, language, artifacts, institutions, connected populations, and recombination; and social systems can perform important allostatic work by redistributing energy, information, labor, and risk across individuals and time.
We remain genuinely unsure about: the historical order and relative weight of cooperative care, dietary change, cooking, language, and cumulative culture; how much each contributed to particular changes in hominin anatomy and life history; which features best explain the unusual open-endedness of human culture; how often socially transmitted behavior has redirected genetic evolution in nonhuman species; and whether biological evolution belongs within a deeper physics of adaptive organization.
The secure claims are enough to support the unit. The unsettled claims concern the historical details and the best level at which to describe the same reciprocal loop.
48.5 Social allostasis
Allostasis is usually described within the boundary of one organism. A brain evaluates conditions and reallocates bodily resources — increasing cardiovascular output, releasing stored fuel, suppressing one activity to fund another. Yet social living changes who bears the cost of regulation.
A hungry individual can be fed by another. A sick individual can be protected while unable to forage. Several adults can divide vigilance and childcare. A group can pool food obtained at different times and places, reducing the variance faced by each person. Information about a distant water source or a dangerous animal can spare others the cost of discovering it independently. Regulation is no longer limited to what one body can accomplish with its own current reserves.
This is the strongest sense in which the human social niche is allostatic: it redistributes resources across individuals, places, and time. Food sharing moves energy from a successful forager to an unsuccessful one. Storage moves energy from abundance to shortage. Care moves labor toward an injured person or dependent child. Teaching moves information from the experienced to the inexperienced. Rules and institutions coordinate these transfers so that they need not be renegotiated from nothing on every occasion. A social group can absorb fluctuations that would overwhelm an isolated body.
This does not mean society abolishes allostatic load. Social systems can impose it. Hierarchy, exclusion, violence, exploitation, and unstable obligations can themselves become chronic demands. A group may buffer some members while extracting resources from others. Human institutions are control systems with contested targets: stability for whom, achieved at whose cost? The language of regulation should never be allowed to smuggle harmony into a system structured by conflict.
Still, the biological consequence is clear. Human bodies are regulated through relationships. An individual who appears self-sufficient is usually standing on a dense infrastructure of other people’s labor — food production, sanitation, electricity, transport, medical knowledge, and accumulated technology. The relevant environment is not merely the climate outside the skin. It includes the social arrangements that determine whether calories, care, and information arrive when needed.
The social niche therefore solves some of the same problems as the nervous system, but at another scale. It coordinates resources, buffers disturbance, stores information, and organizes action. It does so imperfectly, without a central controller, and with persistent conflicts among the units involved. Yet it can achieve forms of stability no solitary organism could produce.